Система ЭПХХ, схема подключения эпхх, необходимость система ЭПХХ
просмотров 8 657 Google+Система ЭПХХ предназначена для прекращения подачи топлива при следовании автомобиля в режиме принудительного холостого хода. Этот режим характеризуется частотой вращения коленвала двигателя, который превышает обороты холостого хода при закрытой дроссельной заслонке. Такой режим применяется в городском цикле и следовании под гору в режиме торможения двигателем.
На инжекторных двигателях прекращение подачи топлива осуществляет электронная система управления двигателем, а в карбюраторных двигателях блок управления ЭПХХ.
Из чего состоит система ЭПХХ.
В состав система ЭПХХ входит блок управления, электромагнитный клапан или электромагнитный пневмоклапан, датчик положения дроссельной заслонки. В качестве датчика числа оборотов часто используется прерыватель трамблёра.
Датчиком положения дроссельной заслонки может быть микропереключатель, контакты которого размыкаются при закрытой дроссельной заслонке либо датчик-винт на конце, которого крепится провод соединяющий вывод блока управления с массой при открытой дроссельной заслонке.
Датчиком оборотов в карбюраторном двигателе выступает прерыватель трамблёра.
Прекращение подачи топлива осуществляется электромагнитным клапаном или электропневматическим клапаном, в зависимости от конструкции карбюратора. Электромагнитный клапан устанавливается на карбюраторе и перекрывает, при отсутствии питания на нём, своим сердечником канал холостого хода. Электропневматический клапан устанавливается на корпусе автомобиля в разрыве трубки соединяющей впускной коллектор с модулем экономайзера, при включении отключает экономайзер от коллектора и соединяет его с атмосферой.В результате этого клапан экономайзера прекращает подачу топлива.
Принцип работы система ЭПХХ.
При частоте вращения коленвала более 1100 об/мин. и закрытой дроссельной заслонке, блок управления отключает питание с клапана, что прекращает подачу топлива, что позволяет экономить топливо на 2-3% и снизить токсичность выхлопных газов на 15-30%. Кроме этого система ЭПХХ предотвращает детонацию двигателя при отключении двигателя, то есть детонационное сгорание топлива при выключенном зажигании.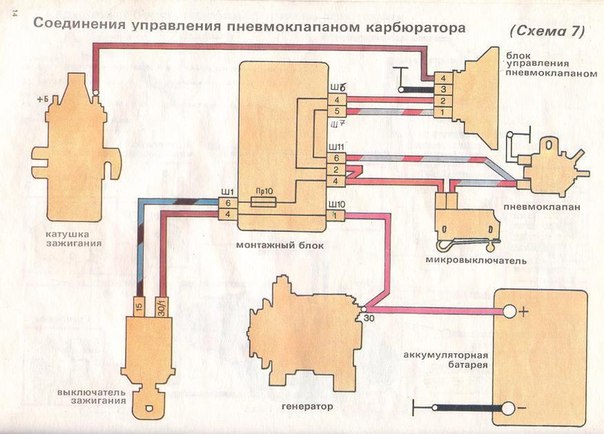
Необходимость системы ЭПХХ.
Езда автомобиля с применением принудительного холостого хода происходит крайне редко, в основном в гористой местности. По этому даже обещанные 2-3% практически не достижимая цель. Но предотвращение детонации двигателя это очень часто необходимо. Но для реализации предотвращения детонации при отключении зажигания не обязательно подключения всей схемы. Для этого достаточно просто подать питание на клапан при включении зажигания и снятие его при выключении.
admin 18/06/2011 «Если Вы заметили ошибку в тексте, пожалуйста выделите это место мышкой и нажмите CTRL+ENTER» «Если статья была Вам полезна, поделитесь ссылкой на неё в соцсетях»Система управления экономайзера принудительного холостого хода
При движении автомобиля в городе, двигатель около 25 % времени работает на принудительном холостом ходу, когда коленчатый вал двигателя вращается за счет кинетической энергии автомобиля и он движется с включенной передачей и отпущенной педалью управления дроссельной заслонкой. В таких режимах двигатель управляется с помощью экономайзера принудительного холостого хода.
В таких режимах двигатель управляется с помощью экономайзера принудительного холостого хода.
Система управления экономайзера принудительного холостого хода УАЗ, блок управления, клапан, микровыключатель.
На принудительном холостом ходу двигатель расходует топливо, не выполняя полезную работу, в результате быстрого закрытия дроссельной заслонки горючая смесь переобогащается и токсичность отработавших газов увеличивается. Для снижения расхода топлива и токсичности отработавших газов на автомобилях УАЗ установлена электрическая система управления экономайзера принудительного холостого хода (ЭПХХ).
Система управления и электрооборудование экономайзера принудительного холостого хода на автомобилях УАЗ с двигателями УМЗ включает в себя блок управления 1422.3733, электромагнитный клапан 1902.3741 и концевой выключатель карбюратора (микровыключатель) 421.3709.
Принцип работы системы управления экономайзера принудительного холостого хода на автомобилях УАЗ.
Режим принудительного холостого хода характеризуется двумя признаками : частота вращения коленчатого вала двигателя больше частоты холостого хода и дроссельная заслонка карбюратора закрыта. Информация о частоте вращения коленчатого вала двигателя поступает в блок управления ЭПХХ с датчика, в качестве которою используется первичная обмотка катушки зажигания, а информация о закрытии дроссельной заслонки — с концевого выключателя, микровыключателя или датчика-винта карбюратора.
Информация о частоте вращения коленчатого вала двигателя поступает в блок управления ЭПХХ с датчика, в качестве которою используется первичная обмотка катушки зажигания, а информация о закрытии дроссельной заслонки — с концевого выключателя, микровыключателя или датчика-винта карбюратора.
Схема соединений электрооборудования экономайзера принудительного холостого хода на автомобилях УАЗ.
При отпускании педали акселератора, в следствии переключения контактов концевого выключателя карбюратора блок управления ЭПХХ вырабатывает сигналы управления электромагнитным (электропневматическим) клапаном подачи топлива в зависимости от частоты вращения коленчатого вала двигателя. Если частота вращения коленчатого вала выше частоты холостого хода, то блок управления снимает напряжение с электроклапана, и подача топлива в двигатель прекращается.
При этом частота вращения коленчатого вала снижается, и когда она станет меньше частоты холостого хода, блок управления подключит напряжение бортовой сети к электроклапану. Подача топлива возобновится и частота вращении коленчатого вала возрастет.
Подача топлива возобновится и частота вращении коленчатого вала возрастет.
При частоте вращения коленчатого вала, снова ставшей больше частоты холостого хода, блок управления опять отключит электроклапан. Процесс повторяется. Периодической отключение подачи топлива в этом режиме снижает расход бензина на 2-3%, а токсичность отработавших газов уменьшается на 15-30%
При нажатии на педаль акселератора контакты концевого выключателя карбюратора переключаются таким образом, что на электроклапан будет постоянно подаваться напряжение бортовой сети. Топливо при этом будет подаваться независимо от частоты вращения коленчатого вала двигателя.
Блок управления 1422.3733 экономайзера принудительного холостого хода на автомобилях УАЗ, принцип работы.
На автомобилях УАЗ с двигателями УМЗ применяются четырехштырьковые блоки управления 1422.3733 экономайзера. В качестве датчик положения дроссельной заслонки используется микровыключатель 421.3709. При закрытой дроссельной заслонке импульсы напряжения пропорциональные частоте вращения коленчатого вала, поступают с первичной обмотки катушки зажигания 1 на вход полупроводникового ключа, собранного на транзисторе VT1.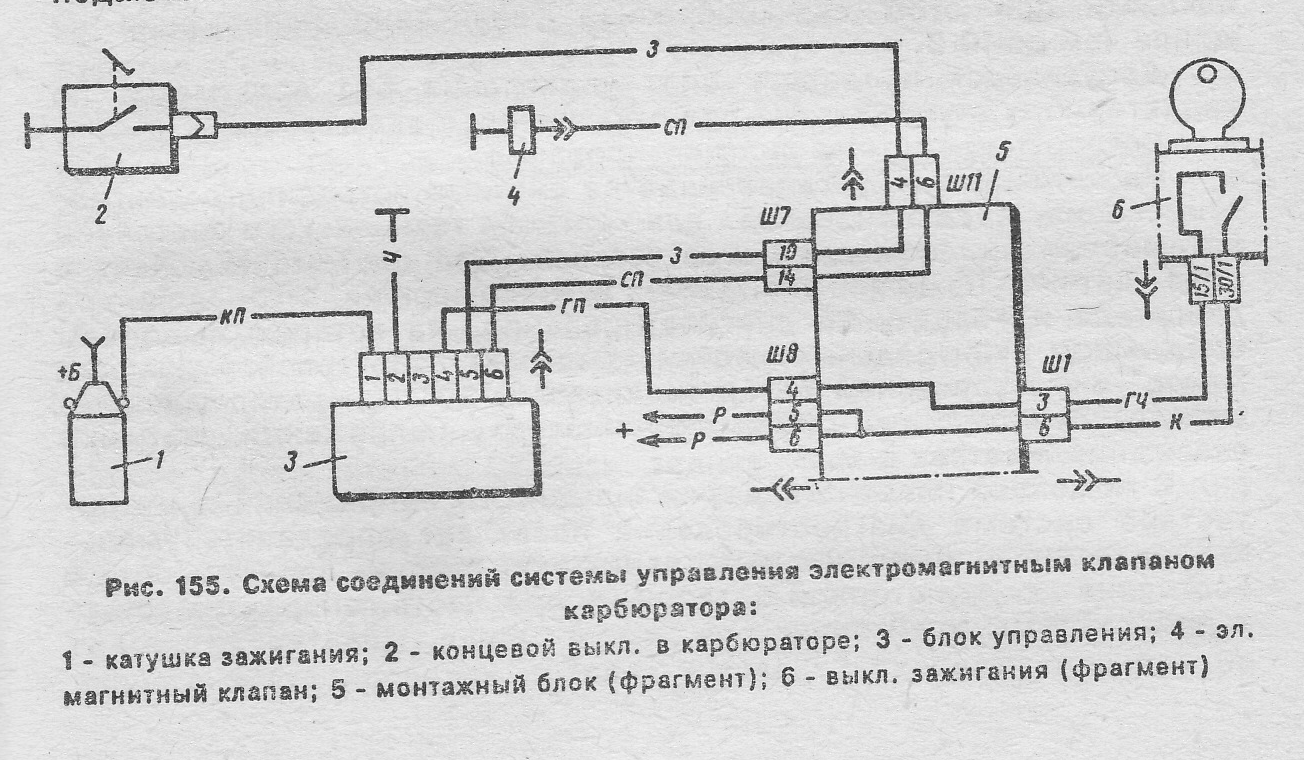
Принципиальная схема блока управления 1422.3733 экономайзера принудительного холостого хода на автомобилях УАЗ с двигателями УМЗ.
Во время действия импульса ключ открывается и конденсатор СЗ разряжается. В паузах между импульсами конденсатор СЗ заряжается. Время заряда, а следовательно, и напряжение на СЗ увеличивается с уменьшением частоты вращения коленчатого вала. При частоте больше частоты холостого хода напряжение на СЗ мало, транзисторы VT2, VT4, VT5, VT6 закрываются. На электромагнитный (электропневматический) клапан напряжение не подается.
Клапан закрывается и подача топлива прекращается. Частота вращения коленчатого вала падает. При частоте меньше частоты холостого хода конденсатор СЗ во время паузы между импульсами успевает зарядиться до напряжения, превышающего опорное напряжение порогового элемента, собранного на транзисторах VT2, VT4. Транзисторы VT2 и VT4 при этом открываются, что обеспечивает открытие транзисторов VT5 и VT6. На электропневмоклапан при этом подается напряжение.
Клапан срабатывает и включает подачу топлива. При открытии дроссельной заслонки контакты микропереключателя S1 замыкаются и напряжение бортовой сети постоянно поступает на электропневмоклапан. Клапан постоянно открыт независимо от сигналов блока управления 1422.3733 экономайзера принудительного холостого хода.
Похожие статьи:
- Регуляторы напряжения РР350 и РР132А, характеристики, устройство и принцип действия.
- Генераторы Г250-Е1 и Г250-П2 на УАЗ-469 и УАЗ-3151, характеристики, устройство и принцип действия.
- Датчики уровня жидкостей и снижения давления в контуре тормозов, устройство, принцип действия, проверка исправности.
- Контактная система зажигания УАЗ, состав и общее устройство, схемы контактной системы зажигания.

- Указатель температуры охлаждающей жидкости 14.3807 и датчик ТМ100, проверка и диагностика неисправностей.
- Указатель давления масла 15.3810 и датчик давления ММ358, проверка исправности, основные характеристики.

Конструкция, проверка и регулировка элементов экономайзера УАЗ-3151, -31512, -31514, -31519
Блок управляет работой электромагнитного клапана в зависимости от частоты вращения коленчатого вала.
На режиме холостого хода блок управления подает напряжение на выводы электромагнитного клапана, который открывается и пропускает разрежение из задроссельного пространства карбюратора к экономайзеру. Клапан экономайзера в свою очередь открывает канал холостого хода.
При повышении частоты вращения свыше 1600 мин–1, блок отключает питание обмотки электромагнитного клапана. При этом пространство над диафрагмой экономайзера через клапан соединяется с атмосферой, разрежение падает, и клапан экономайзера под действием пружины перекрывает канал холостого хода.
Когда частота вращения снижается до 1300 мин–1, блок управления вновь подает напряжение на электромагнитный клапан.
Клапан открывается, пропуская разрежение к диафрагме экономайзера и, клапан экономайзера открывает канал холостого хода.
Электромагнитный клапан обесточивается при выключении зажигания, тем самым исключается самовоспламенение топливно-воздушной смеси в цилиндрах.
Микропереключатель установлен на карбюраторе. При открывании дроссельной заслонки он подает напряжение на электромагнитный клапан, минуя блок управления.
Проверка и замена электромагнитного клапана системы ЭПХХ
Выключаем зажигание и снимаем наконечники проводов с выводов электромагнитного клапана
Снимаем шланги с патрубков клапана, пометив их положение
Крестовой отверткой откручиваем винт крепления клапана и наконечника провода «массы»
Проверяем работу клапана, для чего подаем напряжение 12 В (постоянного тока) на выводы клапана.
Ртом создаем давление в его боковом патрубке.
У исправного клапана воздух должен выходить через второй патрубок, а при снятии напряжения — через воздушный фильтр клапана.
Неисправный клапан заменяем.
Устанавливая клапан, убедитесь в правильности подсоединения шлангов к нему: боковой патрубок соединяется с экономайзером, а центральный — с задроссельным пространством карбюратора.
Подсоединяем наконечники проводов к выводам клапана в произвольном порядке.
Проверка и замена блока управления
Проверку блока управления ЭПХХ рекомендуется проводить автомобильным тестером с функцией тахометра
Отсоединяем провода от микропереключателя карбюратора
Снимаем с наконечников проводов изоляционные трубки
Подсоединяем к выводам электромагнитного клапана зажимы вольтметра
Вместо вольтметра можно использовать контрольную лампу
Подсоединяем тестер (тахометр) к двигателю и пускаем двигатель
Вольтметр должен показать наличие напряжения
Увеличиваем частоту вращения коленчатого вала двигателя.
Когда частота достигнет 1600 мин–1, блок должен отключить подачу электропитания на клапан, а после падения частоты вращения до 1200 мин–1 — вновь включить.
Неисправный блок питания заменяем.
При выключенном зажигании, под панелью приборов, отсоединяем колодку проводов от блока управления
Крестовой отверткой откручиваем два винта крепления блока
Устанавливаем новый блок в обратном порядке
Регулировка положения микропереключателя
Отсоединяем наконечник провода от микропереключателя.
Подсоединяем к его выводам омметр.
При отпущенной педали «газа» контакты микропереключателя должны быть разомкнуты (омметр должен показывать большое сопротивление), при легком нажатии на педаль — замкнуты (сопротивление, показываемое омметром, близко к нулю).
Для регулировки отверткой ослабляем два винта крепления микропереключателя
Перемещая микропереключатель, добиваемся его оптимального положения, после чего затягиваем винты
Эпхх расшифровка.
 Система управления экономайзера принудительного холостого хода на УАЗ, принцип работы блока управления, клапана, микровыключателя
Система управления экономайзера принудительного холостого хода на УАЗ, принцип работы блока управления, клапана, микровыключателяДля управления клапаном ЭПХХ в карбюраторных двигателях автомобилей ВАЗ 2108 — 2110 используется блок управления ЭПХХ 50.3761. В качестве датчика положения дроссельной заслонки используется датчик-винт, представляющий из себя пластмассовый винт с металлическим наконечником, вкручивающийся в кронштейн, закреплённый на карбюраторе.
При открытии дроссельной заслонке наконечник винта, с прикреплённым к нему проводом, не упирается в рычаг дроссельной заслони. Это приводит к разрыву цепи вывода 5 блока управления с массой. При этом закрывается транзистор VT7, а транзистор VT5 открывается, открывая в свою очередь транзистор VT8. Транзистор VT8 подаёт питание на электромагнитный клапан независимо от числа оборотов коленвала.
На вывод 3 блока управления подходит провод, соединяющий его с выводом первичной катушки зажигания, передающий импульсы, которые поступают на вывод 4 микросхемы А1.
Схема блока управления ЭПХХ 25.3761 отличается в основном только работой при оборотах коленчатого вала более 1100 об./мин. Это обусловлено применением в качестве датчика положения дроссельной заслонки микропереключателя, подающего питание на электропневматический клапан при открытой заслонке. Работа блока управления на холостом ходу идентична блоку 50.3761.
Блок ЭПХХ 1402.3733.
Блок ЭПХХ 1402.3733 устанавливается на автомобили семейства ГАЗ и УАЗ. Принцип его работы такая же как и блока 50,3761. Отличие блоков только в схеме.
Принцип его работы такая же как и блока 50,3761. Отличие блоков только в схеме.
Неисправность блока управления ЭПХХ.
При неисправности блока ЭПХХ двигатель не будет работать на холостом ходу или при отпущенной педали газа, обороты скачут от 900 до 1200. Для поиска неисправности достаточно просто удалить сердечник на клапане или соединить трубки на карбюраторе помимо «баллончика».
Система ЭПХХ предназначена для прекращения подачи топлива при следовании автомобиля в режиме принудительного холостого хода. Этот режим характеризуется частотой вращения коленвала двигателя, который превышает обороты холостого хода при закрытой дроссельной заслонке. Такой режим применяется в городском цикле и следовании под гору в режиме торможения двигателем. На инжекторных двигателях прекращение подачи топлива осуществляет электронная система управления двигателем, а в карбюраторных двигателях блок управления ЭПХХ.Из чего состоит система ЭПХХ.
В состав система ЭПХХ входит блок управления, электромагнитный клапан или электромагнитный пневмоклапан, датчик положения дроссельной заслонки. В качестве датчика числа оборотов часто используется прерыватель трамблёра.
В качестве датчика числа оборотов часто используется прерыватель трамблёра.
Датчиком положения дроссельной заслонки может быть микропереключатель, контакты которого размыкаются при закрытой дроссельной заслонке либо датчик-винт на конце, которого крепится провод соединяющий вывод блока управления с массой при открытой дроссельной заслонке.
Датчиком оборотов в карбюраторном двигателе выступает прерыватель трамблёра.
Прекращение подачи топлива осуществляется электромагнитным клапаном или электропневматическим клапаном, в зависимости от конструкции карбюратора. Электромагнитный клапан устанавливается на карбюраторе и перекрывает, при отсутствии питания на нём, своим сердечником канал холостого хода. Электропневматический клапан устанавливается на корпусе автомобиля в разрыве трубки соединяющей впускной коллектор с модулем экономайзера, при включении отключает экономайзер от коллектора и соединяет его с атмосферой.В результате этого клапан экономайзера прекращает подачу топлива.
Принцип работы система ЭПХХ.
При частоте вращения коленвала более 1100 об/мин. и закрытой дроссельной заслонке, блок управления отключает питание с клапана, что прекращает подачу топлива, что позволяет экономить топливо на 2-3% и снизить токсичность выхлопных газов на 15-30%. Кроме этого система ЭПХХ предотвращает детонацию двигателя при отключении двигателя, то есть детонационное сгорание топлива при выключенном зажигании.
Необходимость системы ЭПХХ.
Езда автомобиля с применением принудительного холостого хода происходит крайне редко, в основном в гористой местности. По этому даже обещанные 2-3% практически не достижимая цель. Но предотвращение детонации двигателя это очень часто необходимо. Но для реализации предотвращения детонации при отключении зажигания не обязательно подключения всей схемы. Для этого достаточно просто подать питание на клапан при включении зажигания и снятие его при выключении.
Экономайзер принудительного холостого хода или ЭПХХ позволяет в значительной мере уменьшить выброс токсических веществ в атмосферу. Также он понижает потребление топлива.
Также он понижает потребление топлива.
Что собой представляет экономайзер
Устройство и схема подключения экономайзера
- катушка зажигания,
- изолированный наконечник,
- винт,
- электромагнитный клапан,
- блок управления ЭПХХ.
Каждая из этих деталей взаимодействует друг с другом. Результатом подобного процесса является повышенная производительность мотора и существенное увеличение экономии топлива. Но для того чтобы достигнуть такого результата всё должно быть правильно подключено. О том, как это сделать вы можете узнать из схемы подключения ЭПХХ ниже.
Принцип работы
Есть такое понятие, как торможение двигателем. Проще говоря, это ситуация, когда автомобиль продолжает своё движение по инерции. При этом передача всё ещё включена, а педаль, отвечающая за карбюратор отпущена. Подобное состояние также называется принудительным холостым ходом. Отсюда, собственно, и аббревиатура.
Подобное состояние также называется принудительным холостым ходом. Отсюда, собственно, и аббревиатура.
При этом внутри двигателя происходят очень интересные и важные процессы. Естественно, топливная смесь в цилиндрах продолжает воспламеняться. Но при этом эффективность работы системы падает в несколько раз. Как результат отработанные газы имеют повышенное содержание оксида углерода и углеводородов.
Внимание! На принудительном холостом ходу топливо расходуется крайне неэкономно.
Естественно, автомобильные инженеры не могли просто так оставить подобный дефект. Результатом долгих исследований и экспериментов стало изобретение системы ЭПХХ. Она позволяет отключить подачу топлива при работе в режиме холостого хода, тем самым решив ряд описанных выше проблем.
Отключение подачи топлива стало возможным благодаря электромагнитному клапану, который монтируется в крышке карбюратора. В данной конструкции за подачу тока отвечает блок управления. Он вместе с клапаном создаёт одну электрическую цепь, в которую также входит:
- источник питания;
- датчик, фиксирующий положение дроссельной заслонки;
- катушка зажигания,
- масса.

Информация передаётся посредством электрического импульса, который идёт от катушки зажигания. Обычно он содержит данные о частоте вращения. О том, что карбюратор перешёл в режим холостого хода сигнализирует датчик. Это третий контакт, который подключается к одному из винтов. Замыкание делается на массу.
Система ЭПХХ работает таким образом, что на холостом ходу обмотка пятого электромагнитного клапана обесточивается. Результатом этого действия является прекращение подачи топлива.
Для того чтобы подача топлива возобновилась система ЭПХХ при помощи второго блока должна зарегистрировать два изменения:
- Частота вращения коленвала должна превысить отметку в 2000 оборотов за минуту.
- Дроссельная заслонка должна находиться в закрытом положении.
Только тогда, когда эти два условия будут выполнены, система ЭПХХ сможет возобновить подачу топлива. Но не всё так просто. Если с пониманием внутренних процессов никаких сложностей возникнуть не должно, то возникает другой закономерный вопрос, а что для этого нужно сделать водителю?
В действительности всё довольно просто. Чтобы система ЭПХХ возобновила подачу топлива водителю нужно совершить некоторые действия. Вначале необходимо уменьшить скорость движения
. При этом нельзя нажимать на педаль, которая контролирует положение дроссельной заслонки.
Чтобы система ЭПХХ возобновила подачу топлива водителю нужно совершить некоторые действия. Вначале необходимо уменьшить скорость движения
. При этом нельзя нажимать на педаль, которая контролирует положение дроссельной заслонки.
Есть ещё один способ дезактивировать систему ЭПХХ. Для этого вам также нужно вдавить в пол педаль, отвечающую за дроссельную заслонку. Но частота вращения должна быть высокой. Чтобы этого добиться нужно продолжать движение.
Внимание! Система ЭПХХ включает подачу топлива при 150—200 оборотах в минуту.
Отдельно нужно упомянуть особенности работы электромагнитного клапана. Он обесточивается, когда включается зажигание. Подобная предосторожность позволяет исключить то, что двигатель начнёт свою работу с воспламенения.
Неисправности и диагностика ЭПХХ
Система ЭПХХ не отличается особенной сложностью. Именно этот факт выступает гарантией долгой работы. Но даже эта деталь может выйти из строя при больших нагрузках и длительной эксплуатации автомобиля.
Обычно при выходе из строя системы — двигатель не запускается при отпущенной педали. Он просто глохнет. Начать диагностику нужно с проверки шланга, который соединяет пневматический электрический клапан и клапан ЭПХХ.
Внимание! Двигатель может глохнуть из-за того, что в шланге происходит подсос.
Также нужно при диагностике системы ЭПХХ с большим вниманием отнестись к электрическим контактам. Вы должны проверить надёжность соединений. Довольно часто из строя выходит пневматический электрический клапан. Поэтому очень важно осмотреть и его. Следующими на очереди идут ЭБУ и микропереключатель. Проверку можно проводить только при включённом зажигании и неработающем моторе!
Признаком работоспособности пневматического электроклапана будет характерный щелчок, раздающийся при отсоединении и подключении кабеля. Если же этого нет, то дальнейшую проверку нужно осуществлять при помощи контрольной лампы. Это поможет определить есть ли подача тока. При его отсутствии дальше проверяется ЭБУ и микропереключатель.
Итоги
ЭПХХ позволяет добиться значительной экономии топлива. Это крайне выгодное конструктивное решение, позволяющее при минимальных затратах повысить производительность мотора. Отдельным бонусом идёт снижение токсичности отработанных газов.
При движении автомобиля в городе, двигатель около 25 % времени работает на принудительном холостом ходу, когда коленчатый вал двигателя вращается за счет кинетической энергии автомобиля и он движется с включенной передачей и отпущенной педалью управления дроссельной заслонкой. В таких режимах двигатель управляется с помощью экономайзера принудительного холостого хода.
Система управления экономайзера принудительного холостого хода УАЗ, блок управления, клапан, микровыключатель.
На принудительном холостом ходу двигатель расходует топливо, не выполняя полезную работу, в результате быстрого закрытия дроссельной заслонки горючая смесь переобогащается и токсичность отработавших газов увеличивается. Для снижения расхода топлива и токсичности отработавших газов на автомобилях УАЗ установлена электрическая система управления экономайзера принудительного холостого хода (ЭПХХ).
Система управления и электрооборудование экономайзера принудительного холостого хода на автомобилях УАЗ с двигателями УМЗ включает в себя блок управления 1422.3733, электромагнитный клапан 1902.3741 и концевой выключатель карбюратора (микровыключатель) 421.3709.
Принцип работы системы управления экономайзера принудительного холостого хода на автомобилях УАЗ.
Режим принудительного холостого хода характеризуется двумя признаками: частота вращения коленчатого вала двигателя больше частоты холостого хода и дроссельная заслонка карбюратора закрыта. Информация о частоте вращения коленчатого вала двигателя поступает в блок управления ЭПХХ с датчика, в качестве которою используется первичная обмотка катушки зажигания, а информация о закрытии дроссельной заслонки — с концевого выключателя, микровыключателя или датчика-винта .
При отпускании педали акселератора, в следствии переключения контактов концевого выключателя карбюратора блок управления ЭПХХ вырабатывает сигналы управления электромагнитным (электропневматическим) клапаном подачи топлива в зависимости от частоты вращения коленчатого вала двигателя. Если частота вращения коленчатого вала выше частоты холостого хода, то блок управления снимает напряжение с электроклапана, и подача топлива в двигатель прекращается.
При этом частота вращения коленчатого вала снижается, и когда она станет меньше частоты холостого хода, блок управления подключит напряжение бортовой сети к электроклапану. Подача топлива возобновится и частота вращении коленчатого вала возрастет.
При частоте вращения коленчатого вала, снова ставшей больше частоты холостого хода, блок управления опять отключит электроклапан. Процесс повторяется. Периодической отключение подачи топлива в этом режиме снижает расход бензина на 2-3%, а токсичность отработавших газов уменьшается на 15-30%
При нажатии на педаль акселератора контакты концевого выключателя переключаются таким образом, что на электроклапан будет постоянно подаваться напряжение бортовой сети. Топливо при этом будет подаваться независимо от частоты вращения коленчатого вала двигателя.
Блок управления 1422.3733 экономайзера принудительного холостого хода на автомобилях УАЗ, принцип работы.
На автомобилях УАЗ с двигателями УМЗ применяются четырехштырьковые блоки управления 1422.3733 экономайзера. В качестве датчик положения дроссельной заслонки используется микровыключатель 421.3709. При закрытой дроссельной заслонке импульсы напряжения пропорциональные частоте вращения коленчатого вала, поступают с первичной обмотки катушки зажигания 1 на вход полупроводникового ключа, собранного на транзисторе VT1.
Во время действия импульса ключ открывается и конденсатор СЗ разряжается. В паузах между импульсами конденсатор СЗ заряжается. Время заряда, а следовательно, и напряжение на СЗ увеличивается с уменьшением частоты вращения коленчатого вала. При частоте больше частоты холостого хода напряжение на СЗ мало, транзисторы VT2, VT4, VT5, VT6 закрываются. На электромагнитный (электропневматический) клапан напряжение не подается.
Клапан закрывается и подача топлива прекращается. Частота вращения коленчатого вала падает. При частоте меньше частоты холостого хода конденсатор СЗ во время паузы между импульсами успевает зарядиться до напряжения, превышающего опорное напряжение порогового элемента, собранного на транзисторах VT2, VT4. Транзисторы VT2 и VT4 при этом открываются, что обеспечивает открытие транзисторов VT5 и VT6. На электропневмоклапан при этом подается напряжение.
Клапан срабатывает и включает подачу топлива. При открытии дроссельной заслонки контакты микропереключателя S1 замыкаются и напряжение бортовой сети постоянно поступает на электропневмоклапан. Клапан постоянно открыт независимо от сигналов блока управления 1422.3733 экономайзера принудительного холостого хода.
19023741 Клапан электропневматический ВАЗ,ГАЗ,МОСКВИЧ СОАТЭ — 1902.3741
Клапан электропневматический ВАЗ,ГАЗ,Москвич СОАТЭУстановка — под капотом.
Рабочее положение — горизонтальное.
Метод крепления — на кронштейне.
Применяется в системе экономайзера
Предназначение — работа в системе снижения токсичности выхлопных газов двигателя автомобилей.
— Активная среда — воздух с парами бензина АИ-93 (15%)
— Пропускная способность при разрежении 0,06 МПа – 2,78х10-4м3/с
— Номинальное напряжение питания постоянного тока обмотки 12 В
— Сопротивление обмотки 32 — 42 Ом
— Ток, потребляемый катушкой, 0,375 А (не более)
— Диапазон рабочих температур от -40°С до +80°С
— Ресурс (80%-ный) до отказа при правильной эксплуатации не менее 30000 км пробега а/м
— Габаритные размеры:
  — максимальный диаметр 40 мм
  — максимальная длина 70 мм
— Вес: 0,104 кг.
В упаковке: 40 шт.
Использована информация: ЗАО «СОАТЭ» им.А.М.Мамонова
Наличие товара на складах и в магазинах, а также цена товара указана на 20.08.2021 17:30.Цены и наличие товара во всех магазинах и складах обновляются 1 раз в час. При достаточном количестве товара в нужном вам магазине вы можете купить его без предзаказа.
Интернет-цена — действительна при заказе на сайте или через оператора call-центра по телефону 8-800-600-69-66. При условии достаточного количества товара в момент заказа.Цена в магазинах — розничная цена товара в торговых залах магазинов без предварительного заказа.
Срок перемещения товара с удаленного склада на склад интернет-магазина.
Представленные данные о запчастях на этой странице несут исключительно информационный характер.
Схема подключения эпхх на солекс
Итак согласно книги А.Г. Ходасевич, Т.И. Ходасевич «Справочник по устройству и ремонту электронных приборов автомобилей. Часть 3. Системы автоматического управления экономайзером принудительного холостого хода (САУ ЭПХХ) автомобилей» существует три модификации шестиштырькового Блока управления ЭПХХ 50.3761 (далее по тексту БУ ЭПХХ). На самом деле этот блок семиштырьковый, но один контакт не задействован, отсюда путаница в названиях и даже в этой книге.
Как видим для карбюратора «Солекс» существует две модификации, отличающиеся только порогом срабатывания:
1) 50.3761 (503.3761) для ВАЗ-2108 ;-09;-099; ЗАЗ-1102; АЗЛК 21412-01 — порог выключения 2100 об., порог включения 1900 об.;
2) 501.3761 (5013.3761) для ВАЗ с карбюраторами “Солекс” — порог выключения 1900 об., порог включения 1700 об.
Третья модификация блока предназначалась для карбюратора Озон.
Цитата из книги, поясняющая особенности карбюратора «Солекс»:
Исходя из вышеприведенной цитаты пороги включения /отключения ЭПХХ для всех (двух) карбюраторов «Солекс» должен быть одинаков, но из таблицы 1.3 следует, что для «класики» ВАЗ пороги почему-то ниже.
Далее из книги видим, что промышленность выпускала БУ ЭПХХ 50.3761 на разных микросхемах (К425 серии, К554 серии, К561 серии), да еще и с разными выходными транзисторами.
Практика
Вскрытие торцевой крышки с контактами показало, что мой БУ ЭПХХ 50.3761 собран на микросхемах К425 серии, выходной транзистор VT6 КТ427Х.
Ниже скриншоты из книги для этой модификации БУ:
Как видим здесь представлены все три платы модификации БУ ЭПХХ 50.3761. Они отличаются только номиналами резисторов R2*, R5* и R6*. Видимо здесь, в резисторах которые необходимо подбирать, собака и зарыта, то есть разница порогов включения /выключения ЭПХХ.
Номиналы резисторов согласно принципиальной схемы: R2* 20kOm; R5* 4,7kOm и R6* 100kOm (в сумме R5+R6 104,7kOm)
Номиналы резисторов модификации блока вариант 1: R2* 560Om; R5* 3,9kOm и R6* 10МOm (в сумме R5+R6 10,39МOm).
Номиналы резисторов модификации блока вариант 2: R2* 18kOm; R5* 2,7kOm и R6* 110kOm (в сумме R5+R6 112,7kOm).
Номиналы резисторов модификации блока вариант 4: R2* 23kOm; R5* 750Om и R6* 87kOm (в сумме R5+R6 87,75kOm).
Как позже оказалось вариант 4 как раз и соответствует порогам выключения 2100 об. и выключение при 1900 об. Отклонение от параметров резисторов принципиальной схемы и фактических параметров резисторов моего блока объясняются тем, что все радиодетали имеют разброс параметров и изготовитель подбирал параметры R2*, R5* и R6* в каждом конкретном изделии для получения заданных параметров порогов отключения/включения ЭПХХ.
В моем блоке эти резисторы были таких номиналов: R2* 23,2kOm; R5+R6 92,83kOm, т.е. вариант 4. При этом порог выключения 2100 об., порог включения 1900 об.
С помощью припаянных переменных резисторов провел эксперимент: с сопротивлением резисторов как в варианте 2 порог выключения составил 1850 об., а порог включения 1650 об. Вполне можно ездить, двигатель не глохнет, во всяком случае мой с моими доработками карбюратора.
Установка резисторов как в варианте 1 изменила пороги так: порог выключения составил 1100 об., а порог включения 850 об. Естественно двигатель глох при переходе на ХХ.
Вывод
Повышая сопротивление пары резисторов R5+R6 с одновременным снижением сопротивления R2 (в пределах вариантов 4 и 2) можно снизить безболезненно порог выключения с 2100 об. до 1900 об., порог включения с 1900 об. до 1700 об.
Зачем это надо? Ответ находим в этой же книге: «
Опыты показали что на моем двигателе с доработанным карбюратором при пороге выключения 1600 об. и пороге включения 1400 об. при отпускании педали газа с 2500 об. двигатель не глохнет, только происходит кратковременный провал до 650 об. и восстановление до 800 об. ХХ, что допустимо.
Карбюратор 21073 1107010 ДААЗ разрабатывался для автомобилей «Нива» ВАЗ-2121 с объемом двигателя 1,6 л и ВАЗ-21213 с 1,7 литровым двигателем.
Солекс 21073-1107010 является эмульсионным, двухкамерным карбюратором с падающим потоком (движение потока сверху вниз). Дроссельные заслонки открываются механически, последовательно с помощью педали «газа».
Карбюратор имеет следующие узлы и системы:
- Главные дозирующие системы, их две, для первой и второй камер соответственно.
- Поплавковая камера оснащена двойным поплавком, сбалансирована для предотвращения влияния на работу карбюратора наклонов, например при повороте автомобиля.
- Система отсоса картерных газов.
- Механизм, блокирующий открытие дроссельной заслонки второй камеры.
- Система холостого хода связана с первой камерой.
- Экономайзер холостого хода.
- Две переходные системы, по одной для каждой из камер.
- Экономайзер мощностных режимов.
- Ускорительный насос.
- Пусковое устройство.
- Устройство подогрева.
Расположение основных узлов карбюратора показано на рисунках:
Карбюратор состоит из двух половинок, более массивной нижней – корпуса, и верхней – крышки карбюратора. В нижней части карбюратора, в каждой из камер находятся поворотные дроссельные заслонки, управляемые механически. В первой камере в верхней части расположена воздушная заслонка, предназначенная для холодного пуска двигателя. Воздушная заслонка управляется тросом, идущим в салон автомобиля (рычаг подсоса), и вакуумным пусковым устройством.
Через впускной штуцер, топливо, проходя через сетчатый фильтр карбюратора и игольчатый клапан, попадает в поплавковую камеру. Камера состоит из двух секций, сообщающихся между собой, поэтому уровень топлива в них одинаков. Двухсекционная конструкция позволяет уменьшить влияние крена автомобиля на уровень топлива и, как следствие, на работу двигателя.
Эмульсионная трубка с воздушным жиклером
По мере наполнения поплавковой камеры, поплавок, поджимая вверх иглу клапана, перекрывает поступление топлива, таким образом, поддерживает постоянный уровень горючего в карбюраторе.
Из поплавковой камеры топливо через главные топливные жиклеры подается в эмульсионные колодцы, туда же через отверстия в верхней части эмульсионных трубок (воздушные жиклеры) поступает воздух. В колодцах при смешивании топлива и воздуха образуется эмульсия, которая попадает в малые и большие диффузоры карбюратора. Это главная дозирующая система карбюратора.
На разных режимах двигателя, в работу включаются те или иные системы карбюратора.
Работа карбюратора Солекс 21073
При пуске холодного двигателя, для обогащения смеси, в работу вступает пусковое устройство, управляемое из салона автомобиля ручкой подсоса. В максимально вытянутом положении ручка подсоса через тросик привода поворачивает рычаг, полностью закрывая воздушную заслонку (первая камера). При этом дроссельная заслонка первой камеры приоткрывается на размер пускового зазора, который можно настроить регулировочным винтом приоткрывания дроссельной заслонки первой камеры на рычаге.
Пусковое устройство состоит из полости, сообщающейся каналом с пространством впускного коллектора, диафрагмы и штока связанного с воздушной заслонкой. После пуска двигателя разрежение во впускном коллекторе воздействую на диафрагму и шток открывает воздушную заслонку на величину пускового зазора (регулируется винтом пускового устройства). При возврате рукоятки в нормальное, утопленное положение, пусковые зазоры уменьшаются. Зазоры в промежуточных положениях полностью зависят от геометрии рычага и не нуждаются в регулировке. Дроссельная заслонка второй камеры через систему рычагов, при вытянутом подсосе, блокируется, поэтому при нажатии на газ вторая камера в работе не участвует для исключения провалов двигателя.
Система холостого хода (СХХ) предназначена для питания двигателя на минимальных оборотах, не давая ему заглохнуть, когда нагрузка отсутствует. Топливо поступает в СХХ через главный топливный жиклер первой камеры, далее жиклер холостого хода, смешивается с воздухом поступающим через воздушный жиклер холостого хода, а также из широкой части диффузора первой камеры. Такая система подачи воздуха в СХХ обеспечивает устойчивый переход в данный режим. Полученная эмульсия поступает в первую камеру через отверстие расположенное под дроссельной заслонкой. Канал ведущий к выходному отверстию холостого хода перекрывает винт качества. Частота оборотов двигателя регулируется так называемым винтом качества, который определяет величину зазора дроссельной заслонки камеры номер один в режиме холостого хода.
При плавном нажатии на педаль газа, в работу включается переходная система первой камеры. Ее дроссельная заслонка частично открывается, из щели переходной системы, которая расположена выше заслонки, начинает поступать дополнительное топливо, обогащая смесь. Переходная система первой камеры не допускает провал при переходе из режима холостого хода, при трогании автомобиля.
Переходная система второй камеры устроена аналогично, с той лишь разницей, что обогащает смесь при переходе из режима средних к большим нагрузкам, и ее выходное отверстие круглое. Эта система помогает избежать провалов при движении автомобиля.
При достаточно сильном открытии заслонок в работу вступает экономайзер мощностных режимов. Экономайзер забирает топливо непосредственно из поплавковой камеры и управляется разрежением во впускном коллекторе. При закрытой заслонке разряжение велико, и диафрагма экономайзера не воздействует на шариковый клапан, перекрывающий поток топлива. При открытии заслонки разрежение уменьшается, пружина воздействует на диафрагму, а та на шарик клапана, открывая путь топливу через жиклер экономайзера в эмульсионный колодец, и, минуя главный топливный жиклер, обогащает топливную смесь.
В режиме работы на максимальных нагрузках двигателю требуется дополнительное топливо. Его подачу осуществляет эконостат непосредственно из поплавковой камеры, через систему каналов к распылителю во второй камере.
Ускорительный насос еще один узел карбюратора. Ускорительный насос, обогащает топливную смесь при разгоне автомобиля. Состоит он из рычага, диафрагмы и распылителя. Кулачок насаженный на ось дроссельной заслонки, при ее открытии воздействует на рычаг насоса, а тот на диафрагму, накачивающую топливо через распылитель в первую камеру карбюратора. В устройстве насоса предусмотрены два обратных клапана. Первый находится в канале связывающем поплавковую камеру и полость насоса, и открывается при заполнении последней под действием пружины отводящей диафрагму, подобно поршню шприца. Клапан закрывается при нагнетании топлива в распылитель (при нажатии на педаль газа). Второй клапан расположен в распылителе ускорительного насоса. При нагнетании топлива он открывается, если топливо перестает поступать – перекрывает канал распылителя, предотвращая подсос воздуха и не давая вытекать топливу. Профиль кулачка ускорительного насоса определяет его производительность.
Экономайзера принудительного холостого хода (ЭПХХ)
О системе холостого хода было сказано выше. СХХ карбюратора 21073 оснащена электромагнитным клапаном, являющемся частью экономайзера принудительного холостого хода (ЭПХХ). Этот клапан перекрывает каналы холостого хода и переходной системы первой камеры, и предназначен для прекращения подачи топлива при выключении двигателя, а также в режиме принудительного холостого хода (торможение двигателем), для уменьшения токсичности выхлопных газов и экономии топлива. ЭПХХ состоит из концевого выключателя (смотрите на рисунке карбюратора), электромагнитного клапана и блока управления.
При включении зажигания перед пуском двигателя, когда дроссельная заслонка первичной камеры карбюратора закрыта упорный винт (винт количества) с концевым выключателем замкнут на корпус автомобиля. При этом напряжение подается на электромагнитный клапан и он открывает топливный жиклер системы холостого хода.
При запуске двигателя и его работе на режиме холостого хода электромагнитный клапан получает питание от блока управления. С возрастанием частоты вращения коленчатого вала до 2100 оборотов в минуту (при нажатии на педаль газа происходит разрыв соединения концевого выключателя с корпусом автомобиля), блок управления отключается от управления электромагнитным клапаном, но питание на электромагнитный клапан продолжает поступать, до того момента пока концевой выключатель вновь не замкнется на массу. При резком закрытии дроссельных заслонок (принудительный холостой ход) концевой выключатель замыкается на корпус автомобиля и питание на электромагнитный клапан отключается, а игла клапана перекрывает подачу топливной смеси.
При уменьшении частоты вращения коленчатого вала до 1900 оборотов в минуту вновь включается блок управления и на электромагнитный клапан подается напряжение, открывается топливный жиклер и начинается подача смеси из системы холостого хода.
Данный карбюратор имеет схожую конструкцию со всеми карбюраторами линейки «Солекс» Димитровградского автоагрегатного завода (сокращенно ДААЗ), но и имеет некоторые отличия. Поскольку устанавливается он на двигатели с большим рабочим объемом, то и характеристики его систем изменены. Распылитель ускорительного насоса оснащен только одной трубкой идущей в первую камеру. Сетчатый фильтр извлекается после выкручивания штуцера подачи топлива. Карбюратор 21073-1107010 оснащен системой управления рецеркуляцией отработавших газов через штуцеры запресованные в корпус, которые по каналам соединяются с пространством первой камеры над заслонкой дросселя и под ней.
Из таблицы ниже вы сможете узнать какие жиклеры стоят на Солекс 21073 1107010.
Тарировочные данные 21073-1107010
Полезное видео по теме:
Пришла пара, на моей 21043 менять карюратор, купил Солекс 21053. Заодно купил бесконтакное зажигание. Нашёл в инэте схему подключения блока ЭПХХ. Купил блок 31,3763 производство «Астро»
На обратной стороне кортонки написано что 31.3763 для автомобилей 2108-21099. а вот 312.3763 для Нивы. у них различаются частоты срабатывания. Вот какой ставить мне, пойдёт ли тот которыйя купил и от чего зависит частота срабатывания ?
Карбюратор то у меня не нивский (21073) и не восьмёрошный (2108) а для класических двигателей объёмом 1,5-1,6 (цитата АвтоВАЗа).
цитата из инэта Мнение
Много пришло писем, отвечаю в рассылку:
Александр, прочитал Ваш опус и с Вами согласен. У меня 2104 с 1500 двигателем кпп-5, редуктор 2106. Машина неплохо разгоняется до 100 но после становится вялой, а еще в этом году без видимых причин на трассе разрегулировалась система зажигания — т.е. мошность пропала напрочь. Проблему решил регулировкой распределителя, но факт был неприятный. Поделитесь опытон установки на классику бесконтактной системы от 08-09.
С уважением, Олег Кононов
С установкой на классику бесконтактной системы зажигания от 2108-09 нет никаких затруднений. Для этого, Вам необходимо приобрести в магазине, на радиорынке следующее:
1. Трамблер с бесконтактным датчиком (от «Нивы 1.7)
2. Коммутатор (от ВАЗ 2108-09)
3. Катушку зажигания (от 2108-09)
4. Пучек проводов с разьемами (ее называют «косынка»)
Если у Вас стоит система ЭПХХ то Вам необходимо купить блок управления ЭПХХ от 2108-09.
Все. Берете эл.схему от «зубила», монтаж «косынки», трамблера, катушки и коммутатора у Вас отнимает час-полтора если Вы-курящий человек. :-)))
Я выкинул штатный, и поставил «восьмерочный» блок. У меня карбюратор с пневматическим клапаном. Один контакт «микрика» посадил на массу, второй- в схему. Завел, все прекрасно работало часа два. Затем пропали «холостые». Разобрал блок экономайзера. Вылетел мааленький ключевой транзистор, управляющий пневмоклапаном. Змаенил его на КТ818 с пластинчатым радиатором. Все. Езжу не нарадуюсь. Пропали «провалы» присущие штатной системе ЭПХХ.
Экономайзер карбюратора ВАЗ — что это и как работает ЭПХХ
Большинство карбюраторов (кроме совсем древних моделей) применяемых на автомобилях ВАЗ, оснащены двумя типа экономайзеров. Прочитав статью, вы узнаете:
- для чего нужны эти устройства;
- как они работают;
- по каким признакам определяют их состояние;
- как настраивают.
Что такое экономайзер и для чего он нужен
Экономайзером называю устройство, которое регулирует подачу топлива. В карбюраторах автомобилей ВАЗ применяют следующие типы экономайзеров:
- Экономайзер принудительного холостого хода (ЭПХХ), который иногда называют электромагнитным клапаном (ЭМК).
- Экономайзер мощностных режимов (ЭМР).
ЭПХХ установлен в верхней части карбюратора, под воздушным фильтром и состоит из соленоида, пластикового привода (по своим функциям аналогичен игольчатому клапану) и жиклера холостого хода. Он отключает подачу топлива по каналу холостого хода в смесительную камеру, если выполнены два условия – скорость вращения коленчатого вала превышает 1,7–2 тысячи оборотов в минуту и нога водителя не находится на педали газа. Сигнал на включение ЭПХХ подает блок управления, подключенный к микровыключателю и системе зажигания. ЭХПП серьезно экономит топливо при движении в горной местности. Во время затяжных спусков он блокирует подачу топлива по системе холостого хода и автомобиль переходит в режим торможения двигателем. Кроме экономии топлива это повышает безопасность движения, ведь на затяжном спуске управляемость и устойчивость автомобиля на пониженной передаче гораздо выше, чем на нейтралке.
ЭМР установлен в нижней части карбюратора, под ЭПХХ. Это устройство состоит из подпружиненной мембраны и клапана. В спокойном состоянии (когда мотор выключен), пружина мембраны давит на шарик, продавливает сопротивление его пружины, благодаря чему бензин свободно проходит через этот клапан, проходит по каналу и поступает в распылитель. Когда мотор работает, разряжение, возникающее ниже дроссельной заслонки, ослабляет влияние пружины диафрагмы, в результате чего пружина клапана выжимает шарик и последний перекрывает поступление бензина в топливный канал. Если педаль газа нажата сильней, чем на 2/3, разряжение ниже дроссельной заслонки падает и клапан открывает путь топливу к распылителю смесительной камеры. В результате смесь становится более обогащенной, что обеспечивает увеличение крутящего момента двигателя.
Признаки неисправности экономайзеров
Вот список признаков, которые могут говорить о неисправности одного из экономайзеров:
- неустойчивая работа на холостых оборотах;
- затрудненный пуск прогретого двигателя;
- увеличившийся расход топлива;
- падение мощности и одновременное увеличение расхода топлива;
- капли бензина в районе ЭМР.
Неустойчивая работа двигателя на холостых оборотах может возникнуть из-за неисправности ЭПХХ. При включении зажигания, блок управления подает на клапан напряжение 12 вольт, в результате чего соленоид втягивает пластиковый привод, перекрывающий проход бензина через жиклер холостого хода. Еще одна причина неустойчивой работы на холостых оборотах – грязь в соответствующем жиклере. Пуск прогретого двигателя происходит через систему холостого хода при полностью отпущенной педали газа. Если пуск затруднен и требуется нажатие на педаль газа, скорее всего, забит жиклер или поврежден соленоид.
Видео — Установка системы ЭПХХ
Увеличение расхода топлива может быть связано с множеством факторов, в том числе с неправильной работой ЭМР. Если пружинка клапана ослабла или поломалась, то клапан экономайзера будет открыт постоянно, переобогащая топливовоздушную смесь. При полностью нажатой педали газа это увеличивает мощность двигателя, но в остальных режимах наоборот, приводит к падению мощности. Из-за этого водитель вынужден сильней давить на газ, что еще больше увеличивает расход топлива. Если диафрагма ЭМР потеряла герметичность или плохо затянута крышка, то бензин будет попадать во впускной коллектор ниже дроссельной заслонки, а также просачиваться наружу. Последнее особенно опасно, потому что может привести к возгоранию топлива.
Диагностика и ремонт ЭПХХ
Как проверять основные системы карбюратора, снимать его с впускного коллектора и сливать топливо, читайте в статье (Карбюратор). Также внимательно прочитайте статью (техника безопасности для ремонта и обслуживания автомобилей), это поможет вам избежать возгорания топлива.
Заменить ЭПХХ или прочистить его жиклер можно без снятия карбюратора. Снимите воздушный фильтр, отсоедините провод от экономайзера и выкрутите его из корпуса карбюратора. Снимите жиклер с пластикового привода и промойте. С помощью двух проводков присоединяйте ЭПХХ к аккумулятору, если привод втягивается больше, чем на 5 мм, экономайзер исправен. Если нет, его необходимо заменить. Не забудьте продуть всю систему холостого хода. Для этого прысните в отверстие для установки ЭПХХ очиститель карбюратора и через 1 минуту продуйте с помощью компрессора.
Любые работы, связанные с ЭМР, проводите только на снятом карбюраторе, слив с него топливо. Положите нижнюю часть карбюратора на чистый стол и открутите 3 винта крепления крышки и диафрагмы. Снимите крышку и дифрагму, не потеряйте пружинку. Снимите шланг вакуумного регулятора опережения зажигания и наденьте его на клапан. Попытайтесь всосать воздух через этот шланг. Если клапан исправен, то воздух не пройдет. Если воздух проходит, клапан необходимо заменить.
Завод ДААЗ, основной поставщик карбюраторов для автомобилей ВАЗ, не выпускает запасные клапаны, поэтому его придется либо снимать с другого карбюратора, либо использовать продукцию других производителей. Чтобы снять клапан, потребуются плоская отвертка и паяльная лампа. С помощью паяльной лампы нагрейте нижнюю часть карбюратора до температуры 100–120 градусов и утконосами вытащите клапан из седла. Не перегревайте карбюратор. Когда карбюратор остынет, обязательно прочистите все каналы ЭМР. Перед установкой нового клапана нагрейте карбюратор до 80–90 градусов. Затем вставьте новый клапан и легкими ударами через оправку, внутренний диаметр которой чуть-чуть больше разрезанной трубки клапана, запрессуйте его на место. Когда карбюратор остынет, установите новую мембрану, пружинку и крышку ЭМР. Закрутите винты и соберите карбюратор, после чего установите его на место.
Полиморфизмы генотипов GSTT1, GSTM1 и EPHX у пациентов с криптогенной полинейропатией: исследование случай – контроль
Brain Behav. 2011 ноя; 1 (2): 135–141.
Йонас Линд
1 Отделение неврологии, Отделение внутренней медицины, Окружная больница Рыхов, Йёнчёпинг, Швеция
Петер Сёдерквист
2 Отделение клеточной биологии, Отделение биомедицины и хирургии, Линчёпингский университет, Линчёпинг Швеция
Матс Фредриксон
3 Кафедра медицины труда и окружающей среды, Университетская больница, Линчёпинг, Швеция
Шахрзад Хоссейниния
4 Кафедра медицины труда и окружающей среды, Медицинский факультет Тегеранского университета, Тегеран, Тегеран
Мартин Тондель
5 Медицина труда и окружающей среды, Департамент общественного здравоохранения и общественной медицины, Гетеборгский университет, Гетеборг, Швеция
Бодил Перссон
3 Департамент медицины труда и окружающей среды, Университетская больница, Линчёпинг, Швеция
Магну s Vrethem
6 Отделения неврологии и нейрофизиологии, Университетская клиника Линчёпинга, Линчёпинг, Швеция
1 Отделение неврологии, Отделение внутренней медицины, Окружная больница Рихов, Йёнчёпинг, Швеция
2 Отделение сотовой связи Биология, кафедра биомедицины и хирургии, Университет Линчёпинга, Линчёпинг, Швеция
3 Кафедра медицины труда и окружающей среды, Университетская больница, Линчёпинг, Швеция
4 Кафедра медицины труда и окружающей среды, Медицинский факультет Университета Тегеран, Тегеран, Иран
5 Медицина труда и окружающей среды, Департамент общественного здравоохранения и общественной медицины, Гетеборгский университет, Гетеборг, Швеция
6 Кафедры неврологии и нейрофизиологии, Университетская больница Линчёпинга, Линчёпинг, Швеция
Джонас Линд, Отделение неврологии, Депутат т внутренних болезней, Окружная больница Рыхова, S-551 85 Йёнчёпинг, Швеция.Тел: +4636322020; Факс: +4636322188; Электронная почта: [email protected].Финансируется за счет грантов FORSS (Совет медицинских исследований Юго-Восточной Швеции) и Futurum — академии здравоохранения при совете графства Йёнчёпинг.
Поступило 14.06.2011; Пересмотрено 2 сентября 2011 г .; Принято 12 сентября 2011 г.
Copyright © 2011 Авторы. Опубликовано Wiley Periodicals, Inc. Эта статья цитировалась в других статьях в PMC.Abstract
Целью данного исследования было проанализировать, могут ли полиморфизмы нулевых аллелей глутатион-S-трансферазы Mu-1 (GSTM1), глутатион-S-трансферазы тета-1 (GSTT1) и генетической вариации эпоксида с низкой активностью. экзон три гидролазы (EPHX * 3) влияет на риск развития полинейропатии.Ферменты этих генов играют важную роль в метаболизме токсичных соединений. Семьдесят девять пациентов с криптогенной полинейропатией (эквивалентной хронической идиопатической аксональной нейропатии) и 398 контрольных пациентов были протестированы на генетический полиморфизм. Медицинские карты были изучены для сбора данных, касающихся клинических проявлений при постановке диагноза, а данные о воздействии были собраны с помощью анкет. Отношения шансов (OR) для нулевых форм GSTM1 и GSTT1 и YY-формы нормальной активности EPHX * 3 были близки к единице, за исключением GSTT1, который достиг 1.86. Самый высокий риск полинейропатии был обнаружен у курильщиков с нулевым GSTT1, у которых риск повышался в 3,7 раза. Взаимодействия между генами были проанализированы и подтвердили увеличение OR для GSTT1, которое было самым сильным, если пациенты имели низкоактивную форму HH EPHX * 3 (OR 2.37). Наша гипотеза состоит в том, что нулевой полиморфизм GSTT1 может быть связан с нарушением метаболизма токсичных веществ, что может привести к повреждению нервов в периферической нервной системе.
Ключевые слова: Хроническая идиопатическая аксональная нейропатия, глутатион-S-трансфераза, эпоксидгидролаза, курение, растворители
Введение
Полинейропатия — неврологическое заболевание, которое часто встречается в среднем и позднем взрослом возрасте.Оценки его распространенности варьируются от 2,4% до 8% и зависят от выбора пациентов, то есть от общей популяции или серии больниц, и даже выше у лиц, подвергшихся воздействию различных токсинов, или пациентов с сахарным диабетом (Beghi et al. 1995 ). Хотя существует несколько известных причин полинейропатии, этиология часто остается неизвестной (Martyn and Hughes, 1997), и тогда ее называют криптогенной. Механизмом в этих случаях часто считается воздействие профессиональных факторов или факторов окружающей среды, которые мы исследовали ранее (Tondel et al.2006 г.). Аксонопатия — наиболее частая форма патологии при токсических невропатиях, но основные патологические механизмы неясны. Считается, что нарушение аксонального транспорта приводит к дегенерации уязвимых дистальных нервных сегментов (Spencer et al., 1979; Griffin and Watson 1988), и когда процесс продолжается, дегенерация продолжается проксимально по направлению к телу клетки.
В промышленных условиях и в окружающей среде некоторые соединения нейротоксичны и часто поражают периферические нервы (Spencer and Schaumburg 2000).Одним из примеров является н-гексан, который накапливается в нервной ткани во время хронического воздействия (Feldman 1999). Известно, что он вызывает первичную дегенерацию аксонов при вторичной демиелинизации (Chang et al. 1993).
Биотрансформация экзогенных и эндогенных соединений может играть роль в индивидуальной восприимчивости из-за генетической изменчивости ферментов, участвующих в детоксикации. Метаболизм биотрансформации можно разделить на две фазы. В фазе 1 исходная чужеродная молекула изменяется путем добавления функциональной группы, которая затем может быть сопряжена в фазе 2.Затем конъюгированная молекула может выводиться из организма (Timbrell 2000). Обычно эти шаги приводят к менее токсичной молекуле, но в некоторых случаях происходит обратное.
Эпоксидгидролазы (EPHX) представляют собой пример ферментной системы фазы 1, которая действует путем добавления воды к эпоксиду (Timbrell 2000). Эти ферменты играют важную роль в метаболизме экзогенных химических веществ, таких как полициклические ароматические углеводороды (ПАУ) (Omiecinski et al. 1993). Эпоксиды могут быть частично детоксифицированы микросомальным EPHX (mEPHX), который катализирует их гидролиз с образованием соответствующих дигидродиолов (Oesch 1973).Хотя этот гидролиз обычно считается реакцией детоксикации, поскольку образуются менее токсичные химические вещества, некоторые дигидродиолы, образующиеся из ПАУ, являются субстратами для дополнительных метаболических изменений высокотоксичных, мутагенных и канцерогенных эпоксидов полициклических углеводородных диолов. Таким образом, EPHX * 3 играет ту же двойную роль в детоксикации и активации проканцерогенов, что и в некоторых цитохромах P450 (Benhamou et al. 1998), и, как следствие, может также играть важную роль в нейротоксичности (Guengerich 1982) и в лекарственном препарате. -связанные побочные эффекты.Два аминокислотных полиморфизма были идентифицированы в кодирующей области экзона три (EPHX * 3), обмена тирозина 113 гистидина (Y113H), что приводит к низкоактивной форме фермента (Hassett et al. 1994), которая может влиять на эпоксид. дезактивация в клетке. Пациенты с наследственной оптической невропатией Лебера, гомозиготные по гистидину 113, заболевали раньше, чем пациенты без этого генотипа (Ishikawa et al. 2005). Полиморфизм в экзоне четыре, гистидин 139 аргинин (h239R, rs2234922), был предложен как высокоактивная изоформа mEPHX (Smith and Harrison 1997; Benhamou et al.1998).
Глутатион-S-трансферазы (GST) представляют собой семейство ферментов фазы 2, ответственных за метаболизм широкого спектра ксенобиотиков и канцерогенов (Mannervik 1985). Эти ферменты катализируют конъюгацию глутатиона с широким спектром органических соединений с образованием тиоэфиров, реакцию, которая иногда является стадией процесса детоксикации, приводящей к образованию меркаптуровой кислоты, классического продукта экскреции ксенобиотиков (Mannervik 1985). Было показано, что ферменты GST защищают организмы от повреждения реактивными кислородными соединениями благодаря их способности связываться с глутатионом (Hayes and Strange 2000).На основании сходства аминокислотных последовательностей и перекрестной реактивности антител GST делятся на несколько классов, включая мю и тета. Глутатион-S-трансфераза Mu-1 (GSTM1) и глутатион-S-трансфераза Theta-1 (GSTT1) являются полиморфными у человека, и делеции в генах приводят к фактическому отсутствию активности фермента, особенно с одновременными делециями в генах GSTM1 и GSTT1 ( Абу-Амеро и др., 2009). Генетические вариации могут изменить восприимчивость человека к канцерогенным веществам и токсинам, а также повлиять на токсичность и эффективность некоторых лекарств (Ginsberg et al.2009 г.). Гены, кодирующие класс ферментов mu, как известно, очень полиморфны (Xu et al. 1998). Они участвуют в детоксикации электрофильных соединений, включая канцерогены, терапевтические препараты, токсины окружающей среды и продукты окислительного стресса, путем конъюгации с глутатионом. Сообщалось, что люди с нулевым генотипом GSTM1 и высоким уровнем воздействия растворителей подвергаются повышенному риску развития хронической токсической энцефалопатии, вызванной растворителем (Söderkvist et al., 1996), и болезни Паркинсона (Dick et al.2007). Ген GSTT1 расположен на хромосоме 22. Как для GSTT1, так и для GSTM1 нулевой генотип был связан с повышенным риском оптических невропатий (Abu-Amero et al. 2009) и побочных эффектов лекарств, включая когнитивные нарушения после лечения у пациентов. с медуллобластомой (Barahmani et al. 2009), но не с наследственной оптической невропатией Лебера (Ishikawa et al. 2005) или невропатией у пациентов, получающих химиотерапию на основе оксалиплатина (Lecomte et al. 2006).
Поскольку активность этих ферментов, метаболизирующих ксенобиотики, как правило, необходима для обеспечения эффективной детоксикации, тем самым защищая организм от повреждений, вызванных воздействием, мы проанализировали наличие полиморфизма нулевых аллелей GSTM1 и GSTT1 и генетической вариации mEPHX (низкая активность) влияют на риск развития полинейропатии.Материал и методы Больница, Йёнчёпинг) в возрасте от 40 до 79 лет на момент постановки диагноза (Lindh et al. 2005). Было получено одобрение комитета по этике для повторной проверки записей и контакта с этими субъектами.Медицинские карты были повторно изучены с использованием заранее определенного протокола исследования, включая симптомы, признаки и лабораторные тесты, для подтверждения правильного диагноза в каждом случае. Пациенты с преимущественно демиелинизирующей нейропатией, наследственной нейропатией или любой другой выявленной причиной нейропатии были исключены (Lindh et al. 2005).
Из первоначальной когорты из 168 пациентов 158 были еще живы, и их попросили принять участие в исследовании. Образцы крови были взяты у 79 пациентов (ответ 50%), которые согласились участвовать.Было 54 мужчины и 25 женщин с полинейропатией (средний возраст 71,0 и 68,5 лет соответственно). 89 пациентов, которые не участвовали, были немного старше (72,5 против 70,2 года), имели более высокую клиническую (1,6 против 1,4) и нейрофизиологическую тяжесть (2,0 против 1,8), но различия не достигли статистической значимости, и наблюдались нет разницы в половом распределении.
Контрольную группу составили 398 человек из популяционной контрольной группы из шведской части исследования болезни Паркинсона тех же генетических полиморфизмов, живущих в той же географической области (Dick et al.2007). Контрольную группу составили 198 мужчин и 200 женщин (средний возраст 67,4 и 67,5 лет соответственно). Средний возраст среди случаев был немного выше, чем в контроле (70,2 против 67,5 лет, P <0,05).
Заполненные анкеты для пациентов и контрольных групп о статусе курения и других воздействиях, включая воздействие растворителей или пестицидов, общую анестезию и питьевую воду из частных колодцев, как описано ранее (Tondel et al. 2006; Dick et al. 2007).
Клиническая тяжесть неврологического состояния оценивалась на основе функционального дефицита.Клиническая тяжесть 1 степени (легкая) определялась как незначительные двигательные и / или сенсорные симптомы без функционального дефицита. Степень 3 определялась как тяжелые симптомы с функциональным дефицитом, включая легкую атаксию или, по крайней мере, некоторую потребность в помощи. Степень 2 или умеренная степень тяжести определялась как те симптомы и нарушения, которые находились между 1 и 3 степенью. Точно так же пациенты считались имеющими нейрофизиологические данные 1 степени (легкие), если нейрография и ЭМГ (электромиография) при постановке диагноза показывали незначительное снижение сложных моторных аксональных потенциалов (CMAP), аксональных потенциалов сенсорных нервов (SNAP) или скорости проводимости (CV) по крайней мере в двух нервах.Нейрофизиологические результаты 3 степени (тяжелые) были определены как потеря сенсорных или моторных реакций по крайней мере в двух нервах, как оценивалось в предыдущем исследовании, а степень 2 (умеренная) — как нейрофизиологические результаты между 1 и 3 уровнями (Lindh et al. 2005). .
Собирали цельную кровь и выделяли лейкоцитарную ДНК с помощью набора для очистки ДНК Wizard Genome (Promega Inc., Мэдисон, Висконсин). Нулевые генотипы GSTM1 и GSTT1 оценивали с помощью мультиплексной полимеразной цепной реакции (ПЦР) с β-глобином в качестве внутреннего контрольного гена для успешной амплификации ПЦР (Arand et al.1996). Полиморфизм аминокислот в гене mEPHX (экзон 3 EPHX1) определяли с помощью анализа PCR-RFLP (полиморфизм длины рестрикционного фрагмента) (Lancaster et al. 1996; Smith and Harrison 1997). Для экзона 3 существует три возможных генотипа: YY, YH и HH. Аллель нормальной активности дикого типа — YY, а генотип низкой активности — HH.
Комитет по этике факультета медицинских наук Университета Линчёпинга одобрил проект.
Статистические методы
Статистический анализ проводился с использованием SPSS версии 15.Поскольку ни контрольная группа, ни пациенты с полинейропатией не были нормально распределены по возрасту, статистический анализ проводился с использованием непараметрического метода; тест Крускала – Уоллиса с последующим U-критерием Манна – Уитни для апостериорного анализа (с использованием поправки Бонферони для множественных анализов). Для категориальных переменных использовался критерий хи-квадрат. Для групп, состоящих менее чем из пяти респондентов, анализ проводился с использованием точного критерия Фишера. Относительный риск выражали как отношение шансов (ОШ) с 95% доверительным интервалом (ДИ).Сравнения считались значимыми, если значения P были <0,05. Полиморфизм анализировали независимо от пола, поскольку гены расположены на аутосомах.
Анализ множественной логистической регрессии использовался для получения оценок OR для генетических факторов с поправкой на следующие заранее заданные ковариаты: возраст, пол, когда-либо употреблявшиеся табак, воздействие растворителей или пестицидов, общая анестезия и питьевая вода из частных колодцев. Взаимодействие между генами анализировали с помощью Stata версии 11.0.
Результаты
Всего 79 случаев с криптогенной полинейропатией и 398 контрольных пациентов были протестированы на генетический полиморфизм в генах GSTM1, GSTT1 и mEPHX. Частоты различных генетических полиморфизмов представлены в. Среди контрольной группы было значительно больше людей с нулевым GSTT1 у женщин, чем у мужчин ( P = 0,04), а гомозиготный вариант HH в mEPHX чаще встречался у мужчин ( P <0,01). Остальные варианты у мужчин и женщин не различались.Статистически значимых различий между случаями и контролем не было ни в одной группе.
Таблица 1
Распределение генетических полиморфизмов у пациентов с криптогенной полинейропатией и контрольной группы
| Общее количество субъектов | Мужчины | Женщины | ||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pnp | Элементы управления | Pnp | Элементы управления | Pnp | Элементы управления | |||||||||||||||||||||||||||
| % | n | % | n | % | n | % | n | % | n | 00% 904040 | 50.6 | 201 | 50,5 | 30 | 55,6 | 100 | 50,5 | 10 | 40,0 | 101 | 50,5 | |||||||||||
| GSTM240 | ||||||||||||||||||||||||||||||||
| GSTM240 | 24 | 44,4 | 98 | 49,5 | 15 | 60,0 | 99 | 49,5 | ||||||||||||||||||||||||
| GSTT1 null | 7 | 8,9 | 613 | 3 | 5,6 | 23 | 11,6 | 4 | 16,0 | 38 | 19,0 | |||||||||||||||||||||
| GSTT1 pos | 72 | 337 | 72 | 337 | 175 | 88,4 | 21 | 84,0 | 162 | 81,0 | ||||||||||||||||||||||
| EPHX * 3 YH | 30 | 38,0 | 142 | 35,7 | 369 | 34,8 | 12 | 48,0 | 73 | 36,5 | ||||||||||||||||||||||
| EPHX * 3 г. 47,42 | 4 | 16,0 | 18 | 9,0 | ||||||||||||||||||||||||||||
OR для нулевых форм GSTM1 и GSTT1 и YY-формы EPHX * 3 были близки к единице для всех полиморфизмов, кроме GSTT1, которая достигла 1,86. Когда мужчин и женщин анализировали отдельно, мы обнаружили, что OR EPHX * 3 YH и HH по сравнению с YY составляло 0,7 у мужчин, тогда как у женщин оно составляло 2,1, почти достигая значимости ().
Таблица 2
Анализ генетических факторов по статусу случай-контроль (случаи vs.контрольные)
| Все испытуемые | Мужчины | Женщины | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Исследуемые группы | OR | P Значение | OR | P Значение | OR | P Значение | |||||||
| GSTM1 | Null | 299 (0,61–1,61) | 1 | 0,82 (0,45–1,50) | 0,54 | 1,53 (0,66–3,57) | 0,40 | ||||||
| GSTT1 | Нулевой | 4 положительный 9024 | 0,16 | 2,23 (0,64–7,74) | 0,31 | 1,23 (0,40–3,80) | 1 | ||||||
| EPHX * 3 | YY | YH | 0,64–7,741,09 | 0,81 (0,42–1,57) | 0,62 | 1.99 (0,80–4,50) | 0,16 | ||||||
| YY | HH | 0,96 (0,45–2,05) | 1 | 0,52 (0,20–1,35) | 0,20 | 9 | 0,12 | ||||||
| YY | YH / HH | 1,06 (0,65–1,71) | 0,90 | 0,71 (0,39–1,30) | 0,29 | 2,13 (0,924–590) 9024–35 | |||||||
Что касается клинических результатов, у 24 пациентов были признаны умеренные симптомы, а у 39 пациентов — тяжелые.Не было обнаружено значительных различий между группами по клинической или нейрофизиологической тяжести на момент постановки диагноза, за исключением тенденции для нулевого GSTM1 иметь более серьезные клинические проявления, чем для положительных случаев GSTM1 (среднее значение 1,55 против 1,31, P = 0,064). Аксональная нейропатия наблюдалась у 41 пациента, а комбинированная аксональная и демиелинизирующая нейропатия — у 19 пациентов. Что касается нейрофизиологических данных, у двух пациентов была чистая моторная нейропатия, у 13 пациентов была чисто сенсорная нейропатия и у 64 пациентов была смешанная сенсомоторная нейропатия.Генетические полиморфизмы не были существенно связаны с этими нейрографическими находками.
Мы также исследовали эффекты различных воздействий. В контрольной группе было 189 (47%) курильщиков или бывших курильщиков по сравнению с 43 (54%) курильщиками среди пациентов с криптогенной полинейропатией. О воздействии растворителей во время работы или в свободное время сообщили 24 (30%) пациентов с криптогенной полинейропатией и 132 (33%) пациентов из контрольной группы. О воздействии пестицидов сообщили восемь (10%) пациентов с криптогенной полинейропатией и 29 (7%) пациентов из контрольной группы.Всего 59 (74%) пациентов с криптогенной полинейропатией и 312 (78%) пациентов контрольной группы подверглись общей анестезии, 51 (65%) пациентов с криптогенной полинейропатией и 29 (71%) пациентов с криптогенной полинейропатией. органы управления имели частное водоснабжение, по крайней мере, какое-то время в своей жизни. OR для криптогенной полинейропатии среди лиц, подвергшихся воздействию, показаны на. GSTT1 null среди курильщиков достиг наивысшего OR (3,72, P = 0,08), а EPHX * 3 HH по сравнению с YY среди подвергшихся воздействию растворителя имел самый низкий OR (0.30, P = 0,14). Логистический регрессионный анализ для различных полиморфизмов, пола, возраста и воздействия не выявил каких-либо искажающих эффектов, за исключением того, что увеличение возраста и мужского пола увеличивало риск криптогенной полинейропатии. Взаимодействия между генами были проанализированы и подтвердили увеличение OR для GSTT1, которое было самым сильным, если у пациентов была HH-форма EPHX * 3 (OR 2.37).
Таблица 3
Эффекты генетических полиморфизмов при различных воздействиях (случаи воздействия и контроль)
| Генетические полиморфизмы | Исследуемые группы | Воздействие | Отношение шансов | P P Значение | Нулевое против положительного | Курение | 1.08 (0,56–2,10) | 0,87 |
|---|---|---|---|---|---|
| GSTT1 | Нуль по сравнению с положительным результатом | Курение | 3,72 (0,85–16,2) | 0,08 | |
| EPHX * 3 | по сравнению сY 1,40 (0,69–2,82) | 0,38 | |||
| HH по сравнению с YY | Курение | 0,57 (0,18–1,82) | 0,44 | ||
| YH / HH | по сравнению сYH / HH14 (0,58–2,22) | 0,74 | |||
| GSTM1 | Нулевой по сравнению с положительным | Растворитель | 0,46 (0,18–1,14) | 0,12 | |
| GSTT1 | по положительному 0,28–6,1) | 1 | |||
| EPHX * 3 | YH по сравнению с YY | Растворитель | 0,72 (0,27–1,91) | 0,63 | |
| 9024H по сравнению с30 (0,06–1,40) | 0,14 | ||||
| YH / HH по сравнению с YY | Растворитель | 0,55 (0,22–1,34) | 0,27 | ||
| GSTM4024 Null | |||||
| GSTM4024 по сравнению с | (0,19–4,94)1 | ||||
| GSTT1 | Нулевое против положительного | Пестициды | 0,25 (0,01–4,51) | 0,39 | |
| Y402 | EPHX40 | EPHX40 .33 (0,24–7,28) | 1 | ||
| HH по сравнению с YY | Пестициды | 0,80 (0,07–9,67) | 1 | ||
| YH402 9024H по сравнению с | YH / H24H1 | 0,23–5,89) | 1 | ||
| GSTM1 | Нулевой или положительный результат | Общая анестезия | 1,09 (0,62–1,90) | 0,78 | |
| GSTT1 | |||||
| GSTT1 | GSTT1 | 05 (0,94–1,17)0,55 | |||
| EPHX * 3 | YH в сравнении с YY | Общая анестезия | 1,10 (0,60–2,11) | 0,76 | |
| H241 | |||||
| Общая анестезия | 0,95 (0,39–2,32) | 1 | |||
| YH / HH в сравнении с YY | Общая анестезия | 1,06 (0,61–1,85) | 0,89 | ||
| 0.78 (0,43–1,43) | 0,45 | ||||
| GSTT1 | Нулевое значение по сравнению с положительным значением | Частное водоснабжение | 1,66 (0,57–4,91) | 0,48 | |
| YPHX240 | |||||
| EPHX240 | 1,11 (0,58–2,14) | 1 | |||
| HH по сравнению с YY | Частное водоснабжение | 1,01 (0,41–2,50) | 1 | ||
| HH | YH по сравнению с YH | 1.09 (0,60–1,97) | 0,88 |
Обсуждение
В этом эпидемиологическом исследовании «случай – контроль» пациентов с криптогенной полинейропатией мы изучили связь нулевого полиморфизма GSTM1 и GSTT1 и нескольких полиморфизмов HH экзона 3 EPHX1. экологическое и химическое воздействие. Хотя мы не обнаружили статистически значимого повышения риска, нулевой генотип GSTT1 был связан с почти двукратным повышением риска полинейропатии.Наша гипотеза заключается в том, что нулевой полиморфизм GSTT1 может быть связан с нарушением метаболизма токсичных веществ и реактивного кислорода, что может привести к повреждению нервов, затрагивая несколько участков вдоль моторных и сенсорных аксонов в периферической нервной системе. Это может привести к атрофии аксонов или набуханию аксонов, что приведет к прогрессирующей дистальной дегенерации аксонов. Миелиновая оболочка может разрушаться одновременно с аксоном. Это может способствовать или прямо приводить к аксональной или комбинированной аксонально-демиелинизирующей нейропатии.
Компоненты сигаретного дыма являются примерами экзогенных субстратов, которые токсичны и, кроме того, подвержены биоактивации и могут как прямо, так и косвенно быть нейротоксичными. Мы обнаружили почти четырехкратное повышение риска полинейропатии у курильщиков без GSTT1, что почти достигло статистической значимости. Teunissen и соавторы сообщили о OR 2,1 для текущего курения у пациентов с хронической идиопатической аксональной полинейропатией (Teunissen et al., 2002), а также было обнаружено, что употребление табака может предрасполагать к более раннему развитию и более серьезным симптомам диабетической невропатии ( Tesfaye et al.2005). Наши данные показывают, что этот риск может быть объяснен курильщиками, несущими определенные генетические полиморфизмы, приводящие к нарушению детоксикации токсичных соединений в сигаретном дыме. В исследовании хронической токсической энцефалопатии, вызванной растворителем, у курильщиков был обнаружен повышенный коэффициент риска 2,5 для нулевого генотипа GSTM1 и 1,5 для нулевого генотипа GSTT1 в общей популяции. У некурящих нулевой генотип GSTM1 не представлял риска хронической токсической энцефалопатии.Ни один из изученных полиморфизмов mEPHX не был связан с повышенным риском. Механизм токсического воздействия сигаретного дыма на нервную систему не известен, но предполагалось, что он опосредован химическими веществами в дыме, наиболее важным компонентом которого считаются ПАУ. Нарушение распада ПАУ у людей с нулевым генотипом GST может привести к усилению воздействия на нервы и, таким образом, к повышенной потере нервов. н-гексан — еще одно токсичное вещество, которое присутствует в сигаретном дыме и, как хорошо известно, вызывает полинейропатию (Zhang et al.2006 г.). Однако в нашем исследовании курение не увеличивало риск полинейропатии. Усиленная отрицательная связь между курением сигарет и активностью GSTM1 была описана при болезни Паркинсона, которая может быть опосредована нейропротективным действием никотина на дофаминергическую систему (De Palma et al. 2010).
Частота нулевого GSTM1 составляет около 42–60% у кавказцев (Garte et al. 2001). Частота гомозиготного нулевого GSTT1 сильно варьируется в зависимости от этнической принадлежности и составляет 10–20% у европеоидов (Rebbeck 1997).Ген EPHX * 3 можно найти в трех различных формах: генотипы дикого типа / с нормальной активностью (YY), гетерозиготные (YH) или гомозиготные / с низкой активностью (HH). В европеоидной популяции около 40% субъектов являются гетерозиготными, а 12% — гомозиготными по генотипу HH (Garte et al. 2001). Частота этих полиморфизмов в нашей исследуемой популяции была аналогичной. Никаких различий в частотах аллелей по возрасту или полу в крупных исследованиях не наблюдалось (Garte et al. 2001). К сожалению, у нас был дисбаланс в нашей контрольной группе: 19% женщин и 12% мужчин имели нулевой полиморфизм GSTT1, а 9% женщин и 18% мужчин имели полиморфизм EPHX * 3 HH.Таким образом, невозможно сделать какие-либо выводы о различиях в рисках криптогенной полинейропатии у мужчин и женщин отдельно.
Лица, несущие гены, кодирующие белки с утраченной или нарушенной функцией, имеют нарушенную метаболическую способность выводить токсичные соединения и, следовательно, могут подвергаться повышенному риску полинейропатии. Этот тип мутации часто имеет OR 2–3 для повышенного риска рака. В нашей группе пациентов с полинейропатией мы обнаружили тенденцию к более низкому OR для гена EPHX * 3 по сравнению с контролем.Общий риск полинейропатии, вероятно, является результатом сложного взаимодействия множества генетических факторов и факторов окружающей среды с течением времени. Ранее мы подтвердили, что производственное воздействие растворителя Стоддарда, выхлопных газов бензина, гербицидов или вибраций рук и ног значительно увеличило риск полинейропатии, а также были выявлены новые детерминанты, то есть диоксид серы, ксилол и метилэтилкетон (Tondel et al. 2006 г.).
Мы не обнаружили какой-либо существенной корреляции между клинической или нейрофизиологической серьезностью и генотипом, за исключением небольшого увеличения тяжести клинических проявлений у пациентов с нулевым GSTM1, которые почти достигли статистической значимости.Возможно, что корреляция может быть обнаружена, если будет использована более чувствительная шкала клинической или нейрофизиологической степени тяжести.
Возможной причиной того, что мы не обнаружили существенных различий, является небольшое количество пациентов. В китайском исследовании 22 случаев полинейропатии на полиграфической фабрике и 163 контрольных случаях была обнаружена ассоциация с CYP2E1 Dra, но не с GSTM1 и GSTT1, что указывает на то, что необходимо либо более выраженное воздействие токсичного агента, либо другие гены могут имеют большее значение для развития полинейропатии (Zhang et al.2006 г.). В выборку нашего исследования вошли пациенты с диагнозом криптогенная полинейропатия в отделениях неврологии. Вполне вероятно, что врачи общей практики правильно диагностировали людей, работающих в промышленных условиях с высоким уровнем воздействия токсичных веществ, или что невропатия поставила им диагноз токсической невропатии, что привело к недооценке риска воздействия в нашем исследовании.
Решением этих проблем могло бы стать проведение Общегеномного исследования ассоциации (GWAS), которое стало успешным способом поиска новых генов-кандидатов, например, при болезни Паркинсона, болезни Альцгеймера (Gandhi and Wood 2010) и спорадический боковой амиотрофический склероз (Шатунов и др.2010). Однако для этого потребуется набрать очень большое количество пациентов из нескольких стран.
В заключение, не было обнаружено значительной корреляции между полиморфизмами GSTM1, GSTT1 и EPHX1 у пациентов с криптогенной полинейропатией по сравнению с контрольной группой. Однако сильная тенденция наблюдалась в отношении нулевого фенотипа GSTT1 и курения у этих пациентов по сравнению с контрольной группой (OR 3,7). Нулевой полиморфизм GSTT1 может быть связан с нарушением метаболизма токсичных веществ и реактивного кислорода, что может привести к повреждению нервов в периферической нервной системе.Это может способствовать или прямо приводить к аксональной или комбинированной аксонально-демиелинизирующей нейропатии.
Благодарности
Это исследование было поддержано грантами FORSS (Совет медицинских исследований Юго-Восточной Швеции) и Futurum — академии здравоохранения Совета графства Йёнчёпинг.
Справочные материалы
- Абу-Амеро К.К., Milcarek B, Bosley TM. Генотипы с делецией GSTM1 и GSTT1 при различных спонтанных оптических невропатиях у арабов. Br. J. Ophthalmol. 2009; 93: 1101–1104.[PubMed] [Google Scholar]
- Arand M, Muhlbauer R, Hengstler J, Jager E, Fuchs J, Winkler L, Oesch F. Протокол мультиплексной полимеразной цепной реакции для одновременного анализа полиморфизмов GSTM1 и GSTT1 глутатион-S-трансферазы . Анальный. Biochem. 1996. 236: 184–186. [PubMed] [Google Scholar]
- Barahmani N, Carpentieri S, Li XN, Wang T, Cao Y, Howe L, Kilburn L, Chintagumpala M, Lau C, Okcu MF. Полиморфизм глутатион-S-трансферазы M1 и T1 может предсказывать побочные эффекты после терапии у детей с медуллобластомой.Neuro. Онкол. 2009; 11: 292–300. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Beghi E, Monticelli ML, (IGPSG) IGPSG Хроническая симметричная симптоматическая полинейропатия у пожилых людей: полевое обследование в двух регионах Италии. I. Распространенность и общие характеристики выборки. Неврология. 1995; 45: 1832–1836. [PubMed] [Google Scholar]
- Бенхаму С., Рейникайнен М., Бушарди С., Дайер П., Хирвонен А. Связь между раком легких и генотипами микросомальной эпоксидгидролазы.Cancer Res. 1998. 58: 5291–5293. [PubMed] [Google Scholar]
- Чанг CM, Yu CW, Fong KY, Leung SY, Tsin TW, Yu YL, Cheung TF, Chan SY. Н-гексановая нейропатия в офсетных принтерах. J. Neurol. Нейрохирург. Психиатрия. 1993; 56: 538–542. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Де Пальма Дж., Дик Ф. Д., Кальцетти С., Скотт Н. В., Прескотт Дж. Дж., Осборн А., Хайтес Н., Моззони П., Негротти А., Скаглиони А. и др. Исследование болезни Паркинсона и употребления табака методом случай-контроль: взаимодействие генов и табака. Mov.Disord. 2010; 25: 912–919. [PubMed] [Google Scholar]
- Дик Ф.Д., Де Пальма Дж., Ахмади А., Осборн А., Скотт Н. В., Прескотт Дж. Дж., Беннет Дж., Семпл С., Дик С., Моццони П. и др. Взаимодействие генов и окружающей среды при паркинсонизме и болезни Паркинсона: исследование Geoparkinson. Ок. Environ. Med. 2007. 64: 673–680. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Feldman RG. н-гексан и метил-н-бутилкетон. В кн .: Профессиональная нейротоксикология и нейротоксикология окружающей среды. Филадельфия, Пенсильвания: Липпинкотт-Рэйвен; 1999 г.С. 290–317. [Google Scholar]
- Ганди С., Вуд Н.В. Полногеномные исследования ассоциации: ключ к разгадке нейродегенерации? Nat. Neurosci. 2010. 13: 789–794. [PubMed] [Google Scholar]
- Гарте С., Гаспари Л., Александри А. К., Амброзон С., Отруп Х., Отруп Дж. Л., Баранова Х., Батум Л., Бенхаму С., Боффетта П. и др. Частоты метаболического полиморфизма генов в контрольных популяциях. Cancer Epidemiol. Биомаркеры Пред. 2001; 10: 1239–1248. [PubMed] [Google Scholar]
- Ginsberg G, Smolenski S, Hattis D, Guyton KZ, Johns DO, Sonawane B.Генетический полиморфизм трансфераз глутатиона (GST): популяционное распределение активности конъюгирования GSTM1, T1 и P1. J. Toxicol. Environ. Здоровье B: крит. Ред. 2009; 12: 389–439. [PubMed] [Google Scholar]
- Гриффин Дж. У., Уотсон Д. Ф. Аксональный транспорт при неврологических заболеваниях. Анна. Neurol. 1988; 23: 3–13. [PubMed] [Google Scholar]
- Guengerich FP. Эпоксидгидролаза: свойства и метаболическая роль. Rev. Biochem. Toxicol. 1982; 4: 4–30. [Google Scholar]
- Hassett C, Aicher L, Sidhu JS, Omiecinski CJ.Человеческая микросомальная эпоксидгидролаза: генетический полиморфизм и функциональная экспрессия in vitro вариантов аминокислот. [Ошибка появляется в Hum Mol Genet 1994 Jul; 3 (7): 1214] Hum. Мол. Genet. 1994; 3: 421–428. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Hayes JD, Strange RC. Полиморфизмы глутатион-S-трансферазы и их биологические последствия. Фармакология. 2000. 61: 154–166. [PubMed] [Google Scholar]
- Ishikawa K, Funayama T., Ohde H, Inagaki Y, Mashima Y. Генетические варианты TP53 и EPHX1 при наследственной оптической невропатии Лебера и их связь с возрастом в начале болезни.Jpn. J. Ophthalmol. 2005. 49: 121–126. [PubMed] [Google Scholar]
- Ланкастер Дж. М., Браунли Х.А., Белл Д.А., Футреал ПА, Маркс Дж. Р., Берчак А., Вайзман Р. У., Тейлор Дж. А. Полиморфизм микросомальной эпоксидгидролазы как фактор риска рака яичников. Мол. Канцерогенный. 1996. 17: 160–162. [PubMed] [Google Scholar]
- Лекомте Т., Ланди Б., Бон П., Лоран-Пуч П., Лорио М.-А. Полиморфизм глутатион-S-трансферазы P1 (Ile105Val) предсказывает кумулятивную невропатию у пациентов, получающих химиотерапию на основе оксалиплатина.Clin. Cancer Res. 2006; 12: 3050–3056. [PubMed] [Google Scholar]
- Линд Дж., Тондель М., Эстерберг А., Вретем М. Криптогенная полинейропатия: клинические и нейрофизиологические данные. Дж. Перифер. Nerv. Syst. 2005; 10: 31–37. [PubMed] [Google Scholar]
- Маннервик Б. Изоферменты глутатионтрансферазы. Adv. Энзимол. Relat. Районы Мол. Биол. 1985; 57: 357–417. [PubMed] [Google Scholar]
- Martyn CN, Hughes RA. Эпидемиология периферической невропатии. J. Neurol. Нейрохирург.Психиатрия. 1997. 62: 310–318. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Оеш Ф. Эпоксидные гидразы млекопитающих: индуцируемые ферменты, катализирующие инактивацию канцерогенных и цитотоксических метаболитов, полученных из ароматических и олефиновых соединений. Xenobiotica. 1973; 3: 305–340. [PubMed] [Google Scholar]
- Omiecinski CJ, Aicher L, Holubkov R, Checkoway H. Периферические лимфоциты человека как индикаторы активности микросомальной эпоксидгидролазы в печени и легких. Фармакогенетика. 1993; 3: 150–158.[PubMed] [Google Scholar]
- Реббек TR. Молекулярная эпидемиология генотипов GSTM1 и GSTT1 глутатион-S-трансферазы человека в отношении предрасположенности к раку. Cancer Epidemiol. Биомаркеры Пред. 1997; 6: 733–743. [PubMed] [Google Scholar]
- Шатунов А., Мок К., Ньюхаус С., Уил М. Е., Смит Б., Вэнс С., Джонсон Л., Велдинк Дж. Х., ван Эс М. А., ван ден Берг Л. Х. и др. Хромосома 9p21 при спорадическом боковом амиотрофическом склерозе в Великобритании и семи других странах: исследование ассоциации на уровне всего генома.Lancet Neurol. 2010; 9: 986–994. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Smith CAD, Harrison DJ. Связь между полиморфизмом гена микросомальной эпоксидгидролазы и восприимчивостью к эмфиземе. Ланцет. 1997; 350: 630–633. [PubMed] [Google Scholar]
- Седерквист П., Ахмади А., Акербак А., Аксельсон О., Флодин Ю. Нулевой генотип M1 глутатион-S-трансферазы как модификатор риска хронической токсической энцефалопатии, вызванной растворителем. Скандинавский журнал работы, окружающей среды и здоровья.1996. 22: 360–363. [PubMed] [Google Scholar]
- Spencer PS, Schaumburg HH. Экспериментальная и клиническая нейротоксикология. Оксфорд: Oxford Univ. Нажмите; 2000. [Google Scholar]
- Spencer PS, Sabri MI, Schaumburg HH, Moore CL. Является ли дефект энергетического обмена нервного волокна причиной дегенерации аксонов при полинейропатиях? Анна. Neurol. 1979; 5: 501–507. [PubMed] [Google Scholar]
- Tesfaye S, Chaturvedi N, Eaton SE, Ward JD, Manes C, Ionescu-Tirgoviste C, Witte DR, Fuller JH.Факторы сосудистого риска и диабетическая невропатия. N. Engl. J. Med. 2005; 352: 341–350. [PubMed] [Google Scholar]
- Teunissen LL, Franssen H, Wokke JH, van der Graaf Y, Linssen WH, Banga JD, Laman DM, Notermans NC. Являются ли сердечно-сосудистые заболевания фактором риска развития аксональной полинейропатии? J. Neurol. Нейрохирург. Психиатрия. 2002; 72: 590–595. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Тимбрелл Дж. Факторы, влияющие на токсические реакции: метаболизм. В: Фрэнсис Т., редактор. Принципы биохимической токсикологии.Лондон: Taylor & Francis Ltd .; 2000. С. 65–110. [Google Scholar]
- Тондель М., Линд Дж., Йонссон П., Вретем М., Перссон Б. Профессиональные детерминанты криптогенной полинейропатии. Нейроэпидемиология. 2006. 26: 187–194. [PubMed] [Google Scholar]
- Сюй С., Ван И, Роу Б., Пирсон В. Р.. Характеристика генного кластера глутатион-S-трансферазы человеческого класса и делеции GSTM1. J. Biol. Chem. 1998. 273: 3517–3527. [PubMed] [Google Scholar]
- Чжан И, Лю Цюй, Дуань Х, Ченг Дж, Цзян С., Хуанг Х, Ленг С., Хэ Ф, Чжэн Ю.Связь между полиморфизмом метаболических генов и восприимчивостью к повреждению периферических нервов у рабочих, подвергшихся воздействию н-гексана: предварительное исследование. Биомаркеры. 2006; 11: 61–69. [PubMed] [Google Scholar]
Полиморфизм Tyr113His микросомальной эпоксидгидролазы: связь с риском рака яичников
Функциональное значение было продемонстрировано in vitro для полиморфизма аминокислотной замены экзона 3 T -> C Tyr113His гена микросомальной эпоксидгидролазы (EPHX).Ранее сообщалось, что более высокая активность или быстрый генотип ТТ связаны с повышенным риском рака яичников, и эта ассоциация может отражать усиленную активацию эндогенных или экзогенных субстратов для более реактивных и мутагенных производных. Компоненты сигаретного дыма являются примерами экзогенных субстратов, подверженных такой биоактивации, и воздействие курения может, таким образом, изменять риск, связанный с полиморфизмом EPHX. Мы изучили 545 случаев эпителиального рака яичников и 287 здоровых контрольных случаев для этого генетического варианта EPHX T-C, чтобы выяснить, был ли генотип TT в австралийской популяции связан с (i) специфическими характеристиками опухоли яичника; (ii) риск рака яичников в целом или для отдельных подгрупп; и (iii) риск рака яичников, особенно у курильщиков.Генотипирование проводили с использованием системы обнаружения последовательностей Perkin-Elmer ABI Prism 7700 для распознавания аллелей флуорогенной полимеразной цепной реакцией. Стратификация случаев рака яичников в соответствии с поведением опухоли (низкий потенциал злокачественности или инвазивность), степенью, стадией и иммуногистохимическим статусом p53 не продемонстрировала какой-либо гетерогенности в отношении генотипа, определяемого полиморфизмом EPHX. Было высказано предположение о гетерогенности в отношении гистологического подтипа (P = 0,03), в основном из-за снижения частоты генотипа TT в эндометриоидных опухолях.Распределение генотипов EPHX не отличалось достоверно между здоровыми контрольными группами и случаями рака яичников (в целом, с низким потенциалом злокачественности или инвазивным) ни в целом, ни после стратификации по статусу курения. Однако генотип TT был связан со сниженным риском инвазивного рака яичников конкретно эндометриоидного подтипа (отношение шансов с поправкой на возраст = 0,38, 95% доверительный интервал = 0,17-0,87). Результаты показывают, что предложенная EPHX-опосредованная биоактивация компонентов сигаретного дыма до мутагенных форм вряд ли будет участвовать в этиологии рака яичников в целом, но что более высокая скорость EPHX-опосредованной детоксикации может снизить риск эндометриоидного рака яичников.Мол. Канцерогенный. 30: 71-78, 2001.
Два экзонных однонуклеотидных полиморфизма в гене микросомальной эпоксидгидролазы совместно связаны с преэклампсией
Письменное одобрение исследования было получено от этического комитета университетской больницы Куопио. Информированное согласие было получено от всех пациентов и 115 контрольных групп и задокументировано.
Информация была собрана ретроспективно в отношении 133 преэкламптических беременностей у первородящих женщин и 115 контрольных женщин без преэклампсии в анамнезе, родивших в университетской больнице Куопио в период с января 1994 г. по декабрь 1998 г.Используя журнал регистрации рождений в Куопио, мы искали пациентов с преэклампсией, перезванивали им, и тогда их просили подписать информированное согласие. В то же время мы собрали образцы крови у контрольных людей, которые родили в той же больнице после неосложненной беременности и имели как минимум две нормальные беременности, включая текущую. При зачислении у контрольных групп брали кровь для анализа ДНК. Для обеспечения однородности генетического фона в это исследование случай-контроль были включены контрольные группы, происходящие из региональной популяции и не имеющие клинических признаков расстройства.
Преэклампсия определялась как развитие гипертонии и впервые возникшая протеинурия (> 300 мг белка в моче за 24 часа) у женщин без протеинурии на исходном уровне. Гипертония была определена в соответствии с действующими руководящими принципами, согласно которым систолическое и диастолическое давление 140 и / или 90 мм рт. Группа по повышенному артериальному давлению при беременности, 2000 г.). 7 Женщины с хронической артериальной гипертензией были исключены из исследования.
ДНК экстрагировали из лимфоцитов периферической крови стандартным методом фенол-хлороформной экстракции. Два варианта последовательности гена EPHX , переход T в C, приводящий к замене тирозина гистидином (Tyr113His) в экзоне 3, и переход A в G, приводящий к замене гистидина аргинином (His139Arg) в экзоне 4, были обнаружены. Последовательность целевой ДНК в экзоне 3 гена EPHX амплифицировали с использованием праймеров 5′-GGG GTC CTG AAT TTT GCT CC-3 ‘и 5′-CAA TCT TAG TCT TGA AGT GAC GGT-3’, а также в экзоне 4 гена EPHX . использованными праймерами были 5′-TCT GGT GCC AGA GCC TGA CCG TGC-3 ‘и 5′-ATG GAA CCT CTA GCA GCC CCG TAC C-3’.Инкубационная смесь содержала 50 нг геномной ДНК, 10 пмоль праймеров для амплификации экзонов 3 или 4, 5 нмоль каждого из четырех дезоксинуклеотидтрифосфатов, 1 ед. ДНК-полимеразы AmpliTaq Gold ™ (Perkin Elmer), 10 мМ трис-HCl ( pH 8,3 при 25 ° C), 50 мМ KCl, 1,5 мМ MgCl 2 в реакционном объеме 25 мкл. Протокол циклов включал начальную денатурацию при 95 ° C в течение 7 минут, затем цикл 95 ° C в течение 1 минуты, 55 ° C в течение 1 минуты (для экзона 3) или 64 ° C в течение 1 минуты (для экзона 4), и 72 ° C в течение 1 мин.Всего было выполнено 35 циклов с окончательным удлинением при 72 ° C в течение 10 мин. Для обнаружения полиморфизма Tyr113His смеси ПЦР, содержащие амплифицированный участок ДНК длиной 198 п.н., инкубировали с 5 ед. Фермента Tth 111 I (Promega), 1 × буфером B (6 мМ Tris-HCl, pH 7,5 при 37 ° С). C, 50 мМ NaCl, 6 мМ MgCl 2 , 1 мМ дитиотреитол) и 1 мкг ацетилированного бычьего сывороточного альбумина в общем объеме 30 мкл в течение 2 ч при 65 ° C. Полиморфизм His139Arg в экзоне 4 был обнаружен путем инкубации смесей ПЦР, содержащих продукт ПЦР длиной 322 п.о., с 5 ед. Rsa I (Promega), 1 × буфер C (10 мМ Tris-HCl, pH 7.9 при 37 ° C, 50 мМ NaCl, 10 мМ MgCl 2 , 1 мМ дитиотреитол) и 1 мкг ацетилированного бычьего сывороточного альбумина в общем объеме 30 мкл в течение 2 часов при 37 ° C. Реакционные смеси подвергали электрофорезу в 3% агарозных гелях и фотографировали в УФ-трансиллюминаторе после окрашивания бромидом этидия.
Двусторонние объединенные t -тесты использовались для анализа непрерывных переменных. Статистический анализ для сравнения частот одноточечных аллелей и генотипов проводился с использованием теста Пирсона χ 2 (двусторонние асимптотические значения P ) с SPSS 9.0 программное обеспечение. Отношения шансов (OR) как оценки относительного риска заболевания были рассчитаны с использованием 95% доверительных интервалов (CI). Мы использовали алгоритм ожидания-максимизации (EM) для получения оценок максимального правдоподобия EPHX частот экзонов 3 и 4 со стандартными отклонениями (SD) (каждый гаплотип, состоящий из пары SNP и неизвестной гаметической фазы; Arlequin ver. 2.000 программного обеспечения). Сравнение частоты гаплотипов между преэкламптической и контрольной группами с абсолютным числом хромосом проводилось с использованием точного критерия Фишера (двусторонние значения P с оценкой Монте-Карло на уровне достоверности 99%).Распределение генотипов по Харди – Вайнбергу в преэкламптической и контрольной группах, а также анализ парного неравновесия по сцеплению (LD) оценивали с помощью Arlequin ver. 2.000 программного обеспечения. Программа RxC, использующая алгоритм Metropolis, использовалась для анализа таблиц сопряженности с целью получения объективных оценок точных значений P со стандартными ошибками (SE) в анализе гаплотипов. Определение мощности проводилось с использованием программного обеспечения nQuery Advisor Release 3.0. Уровень статистической значимости был определен как P <0.05.
Полиморфизм в гене микросомальной эпоксидгидролазы связан с преэклампсией
Преэклампсия представляет собой одно из наиболее серьезных распространенных медицинских осложнений беременности у человека. Однако факторы, стимулирующие начало и прогрессирование болезненного процесса, остаются неуловимыми. Появляется все больше свидетельств того, что дисбаланс между токсичными соединениями, такими как перекиси липидов и свободные радикалы кислорода, и веществами, выводящими токсины и поглощающими вещества, может вносить вклад в патофизиологию преэклампсии.1-3
Многие ферменты, включая цитохромы P450, глутатион S-трансферазы, 5 и эпоксидгидролазы, метаболизируют эндогенные и экзогенные соединения. 6-8 Микросомальный эпоксидгидролаза (EPHX, EC 3.3.2.3) катализирует гидролиз арена и оксидов щелочных металлов с образованием транс -дигидродиолы.9 10 Хотя этот гидролиз обычно приводит к детоксикации, поскольку он дает меньше реакционноспособных и более водорастворимых соединений, некоторые производные дигидродиола, особенно в сочетании с окислительным метаболизмом цитохрома P450, являются субстратами для дополнительного метаболизма, в результате чего образуются более реактивные и мутагенные соединения, которые могут связываться с геномной ДНК. .6 8 11 Генетический полиморфизм был описан в гене человека, кодирующем микросомальную эпоксидгидролазу ( EPHX ). Два из этих полиморфизмов, 113Tyr → His в экзоне 3 и 139His → Arg в экзоне 4, были связаны со снижением или увеличением активности фермента соответственно.12 Вариации активности фермента для EPHX в результате таких полиморфизмов могут приводить к изменению индивидуальная предрасположенность к таким заболеваниям, как преэклампсия и HELLP-синдром. Однако пока неизвестно, какие реактивные промежуточные соединения, образованные EPHX, могут быть биологически вероятными с точки зрения предрасположенности к преэклампсии.В этом исследовании случай-контроль мы исследовали возможную связь между генотипами EPHX и риском преэклампсии и HELLP-синдрома. Кроме того, три предполагаемых фенотипа активности фермента EPHX в результате сочетания обоих полиморфизмов были оценены в соответствии с исследованием Hassett et al 12 функциональной экспрессии вариантных аллелей in vitro по остатку 113 (экзон 3, медленный аллель His113) и по остатку 139 (экзон 4, быстрый аллель Arg139). Субъекты с низкой активностью были гомозиготными по His113 или гетерозиготными по His113 в сочетании с гомозиготностью по His139.Субъекты с промежуточной активностью были гомозиготными по Tyr113 и His139 или гетерозиготными по Tyr113 и His139. Субъекты с высокой активностью были гомозиготными по Arg139 или гетерозиготными по Arg139 в сочетании с гомозиготностью по Tyr113.
Материалы и методы
ПАЦИЕНТОВ
Исследовательская группа состояла из 181 белой голландской женщины с преэклампсией в анамнезе с (n = 96) или без (n = 87) синдромом HELLP, которые были описаны ранее.13 Наблюдательный совет учреждения одобрил экспериментальный протокол.Преэклампсия определялась как возникновение после 20 недель беременности диастолического артериального давления выше 90 мм рт. в течение 24-часового периода сбора мочи). Синдром HELLP был определен как одновременное возникновение количества тромбоцитов менее 100 × 10 9 / л, концентрации аспартатаминотрансферазы и аланинаминотрансферазы сыворотки выше 30 МЕ / л, а также гемолиза, установленного на основании аномального мазка крови.Группа из 151 случайно выбранной небеременной белой голландской женщины без основного диабета или почечного, печеночного, гематологического, ранее существовавшего гипертонического или сердечно-сосудистого заболевания, у которых была нормальная нормальная нормальная беременность (среднее количество беременностей 2, диапазон от 1 до 4 ) служил контролем. Общие популяционные характеристики контрольных групп и случаев в период, когда они были госпитализированы по поводу преэклампсии, приведены в таблице 1.
Таблица 1Характеристики пациентов и контрольных групп в период госпитализации по поводу преэклампсии
АНАЛИЗ МУТАЦИЙ
После получения информированного согласия был проведен забор крови.Кровь собирали венепункцией в стерильные, силиконизированные, вакуумные пробирки с этилендиаминтетрауксусной кислотой (ЭДТА) объемом 4 мл (Becton Dickinson, Meylan cedex, Франция). Сразу после сбора кровь хранили при -20 ° C до анализа. Геномную ДНК выделяли из цельной крови с использованием набора для очистки геномной ДНК Wizard TM в соответствии с инструкциями производителя (Promega, Мэдисон, Висконсин, США).
Были исследованы два отдельных полиморфизма в EPHX , а именно 113Tyr → His в экзоне 3 и 139His → 39Arg в экзоне 4.Сначала образцы ДНК амплифицировали с помощью полимеразной цепной реакции (ПЦР) с использованием модификации метода, описанного Hassett et al . 12 Реакционная смесь ПЦР содержала приблизительно 100 нг геномной ДНК, 50 ммоль / л KCl, 10 ммоль / л. Трис HCl, pH 8,0, 2,0 ммоль / л MgCl 2 , 1,5 ед. Полимеразы Taq и 1 мкмоль / л каждого из специфических праймеров прямой (смысловой) и обратной (антисмысловой) амплификации в общем объеме 50 мкл. ДНК денатурировали при 94 ° C в течение пяти минут. Тридцать циклов амплификации начинались с денатурации при 93 ° C, отжига праймеров при 58 ° C для экзона 3 и 65 ° C для экзона 4 и удлинения при 72 ° C, каждый этап в течение одной минуты.Для обнаружения варианта 113Tyr → His продукт ПЦР расщепляли Tth 111 и затем подвергали электрофорезу в 3% агарозном геле. Аллель 113Tyr был идентифицирован по непереваренной единственной полосе ДНК из 231 пары оснований (п.н.), тогда как кодирующий аллель 113His дал две полосы ДНК по 209 п.н. и 22 п.н. Для обнаружения варианта 139His → Arg продукт ПЦР расщепляли Rsa I и затем прогоняли в 3% агарозном геле. Аллель, кодирующий 139His, был идентифицирован двумя полосами ДНК (295 и 62 п.н.), тогда как аллель 139Arg давал три полосы ДНК после переваривания (174, 121 и 62 п.н.).Компания Pharmacia Biotech (Розендал, Нидерланды) синтезировала все праймеры, и все химические вещества, необходимые для ПЦР, были приобретены у Promega (Мэдисон, Висконсин, США).
СТАТИСТИЧЕСКИЙ АНАЛИЗ
Клинические характеристики между группами сравнивали с использованием U-критерия Манна-Уитни. Статистическая значимость различий для индивидуальных полиморфизмов между различными группами и прогнозируемой активности фермента (высокая, средняя или низкая активность) была проверена с помощью анализа хи-квадрат (χ 2 ) с поправкой Йейтса в таблицах сопряженности 2 × 3.Статистическая значимость была принята как p <0,05. Все статистические анализы были выполнены с помощью пакета статистических программ SPSS (SPSS Inc, Чикаго, США).
Результаты
Средний возраст между случаями и контрольной группой существенно не отличался. Не было значительных различий в сроках беременности, рождении, материнском возрасте или диастолическом артериальном давлении между группами преэклампсиса с или без HELLP-синдрома.
Распределение полиморфных вариантов в экзоне 3 и экзоне 4 гена EPHX у пациентов и контрольной группы суммировано в таблице 2.
Таблица 2Распределение полиморфных вариантов в экзонах 3 и 4 гена EPHX
В контрольной группе кодирующий аллель 113His в экзоне 3, соответствующий низкой активности фермента, встречался чаще, чем кодирующий аллель 113Tyr. Шестьдесят семь (44%) контролей были гомозиготными по полиморфизму экзона 3 (His113 His113) и 60 (40%) были гетерозиготными (Tyr113 His113). Мы обнаружили статистически значимо более высокую частоту быстрого генотипа Tyr113 Tyr113 в экзоне 3 среди женщин с преэклампсией в анамнезе (29%) по сравнению с контрольной группой (16%, отношение шансов (OR) 2.0, 95% доверительный интервал (ДИ) 1,2–3,7) (χ 2 = 9,99, p = 0,0068) при сравнении генотипа Tyr113 Tyr113 с генотипами Tyr113 His113 и His113 His113. Подгруппа женщин с преэклампсией с HELLP в анамнезе по сравнению с группой преэклампсии без HELLP не показала достоверных различий в встречаемости полиморфных вариантов в экзоне 3 (OR 1,4, 95% CI 0,7-2,8) ( χ 2 = 0,83, p = 0,66). Не было значительных различий в показателях полиморфизма экзона 3 между женщинами с преэклампсией с (n = 98) или без (n = 85) задержкой внутриутробного развития плода (ЗВУР, определяемой как масса тела при рождении ниже 10-го центиля).Двадцать девять процентов случаев с IUGR имели генотип Tyr113 Tyr113 по сравнению с 22% случаев без IUGR (OR 1,2, 95% CI 0,6–2,4, p = 0,58).
Только 3% контролей были гомозиготными по полиморфизму экзона 4 (Arg139 Arg139), а 33% из 151 субъекта были гетерозиготными (His139 Arg139), по сравнению с 7% и 37%, соответственно, преэкламптических случаев с или без АД-синдром. Это привело к незначительному OR 2,2 (95% ДИ 0,7-7,4, p = 0,20) при сравнении генотипа Arg139 Arg139 с генотипом His139 Arg139 и His139 His139.Опять же, подгруппа женщин с преэклампсией в анамнезе с HELLP, по сравнению с группой без HELLP, не показала значимых различий в частоте полиморфных вариантов в экзоне 4 (OR 0,9, 95% CI 0,3-3,3, p = 0,85).
Частоты аллелей для экзона 3 и экзона 4 не различались между случаями и контролем (χ 2 = 1,90, p = 0,17 для экзона 3; χ 2 = 2,88, p = 0,10 для экзона 4, соответственно).
Распределение субъектов с прогнозируемой высокой, средней и низкой активностью EPHX составило 16 человек.0, 26,9 и 57,1%, соответственно, среди женщин с преэклампсией и 4,5%, 18,2% и 77,3%, соответственно, среди контрольной группы. Была обнаружена значительная связь между прогнозируемыми уровнями активности ферментов и преэклампсией (p = 0,0009), что привело к более высокому риску при более высоких прогнозируемых уровнях ферментов.
Обсуждение
Дисбаланс между токсичными и детоксицирующими веществами может играть роль в патофизиологии преэклампсии. Эпоксидгидролаза микросом является одним из ферментов, играющих важную роль в большом количестве процессов детоксикации и в метаболизме эндогенных и экзогенных соединений.В этом исследовании случай-контроль мы попытались выяснить, могут ли вариации в скорости генетического полиморфизма в гене EPHX коррелировать с предрасположенностью к преэклампсии с синдромом HELLP или без него.
Значительная разница в частотах Tyr113 Tyr113 генотипа EPHX наблюдалась между преэкламптической и контрольной группами. Hassett и др. 12 показали, что замена аллеля гистидина 113 на тирозин 113 снижает активность EPHX примерно на 40%, тогда как замена гистидина 139 аргинином 139 увеличивает активность фермента на 25%.Наше исследование показывает, что гомозиготный генотип в экзоне 3 гена EPHX , приводящий к Tyr113 Tyr113 и соответствующий высокой активности фермента, связан с повышенной частотой преэклампсии. Кроме того, прогнозируемый высокий уровень активности EPHX, оцененный по полиморфизму в экзонах 3 и 4 вместе, был связан с преэклампсией. Связь между более высокой частотой генотипа Tyr113 Tyr113 с высокой активностью фермента EPHX в преэкламптической группе и риском преэклампсии может отражать усиленную активацию эндогенными или экзогенными субстратами более реактивных мутагенных производных диола у этих пациентов.
До сих пор было выполнено лишь несколько исследований роли EPHX в репродуктивной системе. Ван и др. 14 недавно показали, что полиморфизм в гене EPHX может влиять на предрасположенность к самопроизвольному аборту. Они показали связь между генотипом His113 His113 и повышенным риском самопроизвольного аборта. Напротив, Lancaster et al 15 обнаружили связь между генотипом Tyr113 и Tyr113 в экзоне 3 и раком яичников.Эти различные результаты могут быть вызваны тем, что в большинстве случаев EPHX играет детоксифицирующую роль, не позволяя высокореактивным эпоксидам модифицировать основные клеточные (макро) молекулы. Однако при определенных условиях EPHX может способствовать активации других соединений, что приводит к отравлению, а не к детоксикации. 11 16 17 Более того, EPHX может участвовать в реакциях стероидогенеза, что может объяснить наблюдение связи между медленным генотипом и защитой от рака яичников.15
Полиморфизмы в гене EPHX также связаны с гепатоцеллюлярным раком, эмфиземой и раком легких. 18-20 Эти результаты могут представлять собой пример органоспецифических различий в восприимчивости к заболеванию в результате генетических полиморфизмов, поскольку генотип EPHX с высокой активностью Tyr113 Tyr113 может защищать от самопроизвольного аборта14 и гепатоцеллюлярного рака 18, тогда как восприимчивость к раку яичников 15, раку легких 20 и преэклампсии может повышаться.
Частоты полиморфных вариантов в экзоне 4 гена EPHX не сильно различаются в нескольких исследуемых популяциях.12 14 19 20 Однако сообщалось о заметных вариациях в показателях полиморфизма в экзоне 3 для разных этнических и географических популяций; диапазоны от 26-69% до 22-56% были зарегистрированы для генотипов Tyr113 Tyr113 и Tyr113 His113 соответственно12. 14 15 18-20 Зарегистрированные частоты генетических полиморфизмов в гене EPHX перечислены в таблице 3. Поразительно, что наши данные о полиморфизмах экзона 3 у белых субъектов больше похожи на опубликованные данные в китайской контрольной группе, чем в других белых популяциях. .14 18
Таблица 3Частоты генетических полиморфизмов в гене EPHX в контрольных парах
Эти данные подтверждают гипотезу о том, что генетические вариации генов, кодирующих ферменты, участвующие в метаболизме экзогенных или эндогенных веществ, могут влиять на риск заболеваний беременности, таких как самопроизвольный аборт и преэклампсия.
Первичное повреждение ДНК и генетический полиморфизм CYP1A1, EPHX и GSTM1 у рабочих на заводе по производству графитовых электродов | BMC Public Health
Boffetta P, Jourenkova N, Gustavsson P: Риск рака в результате воздействия полициклических ароматических углеводородов на рабочем месте и в окружающей среде. Контроль причин рака. 1997, 8 (3): 444-472. 10.1023 / А: 1018465507029.
CAS Статья PubMed Google ученый
Moulin JJ, Wild P, Mur JM, Lafontaine M, Lefer M, Mercier-Gallay M, Villemot P, Whebi V, Coulon JP: Риск рака легких, гортани, глотки и ротовой полости при производстве угольных электродов рабочие.Scand J Work Environ Health. 1989, 15 (1): 30-37.
CAS Статья PubMed Google ученый
Dell’Omo M, Muzi G, Marchionna G, Latini L, Carrieri P, Paolemili P, Abbritti G: Профилактические меры снижают воздействие полициклических ароматических углеводородов на установке графитовых электродов. Occup Environ Med. 1998, 55 (6): 401-406.
Артикул PubMed PubMed Central Google ученый
Monarca S, Pasquini R, Sforzolini GS, Viola V, Fagioli F: Применение анализа мутагенности сальмонелл и определение полициклических ароматических углеводородов на рабочих местах, подверженных воздействию нефтяного пека и нефтяного кокса. Int Arch Occup Environ Health. 1982, 49 (3-4): 223-239. 10.1007 / BF00377932.
CAS Статья PubMed Google ученый
Пасквини Р., Монарка С., Сфорцолини Г.С., Конти Р., Фаджиоли Ф .: Мутагены в моче рабочих с угольным электродом.Int Arch Occup Environ Health. 1982, 50 (4): 387-395. 10.1007 / BF00377835.
CAS Статья PubMed Google ученый
Петри Т., Шмид П., Шлаттер С. Воздействие в воздухе полициклических ароматических углеводородов (ПАУ) и выделение с мочой 1-гидроксипирена рабочими угольных анодных заводов. Ann Occup Hyg. 1996, 40 (3): 345-357.
CAS Статья PubMed Google ученый
Тета М.Дж., Отт М.Г., Шнаттер А.Р.: Наблюдение за смертностью населения на предприятиях по производству углеродных продуктов. Br JInd Med. 1987, 44 (5): 344-350.
CAS Google ученый
Мори I: Смертность от рака среди рабочих, производящих искусственные графитовые электроды: результаты 38-летнего наблюдения. Occup Environ Med. 2002, 59 (7): 473-480. 10.1136 / oem.59.7.473.
CAS Статья PubMed PubMed Central Google ученый
Густавссон П., Белландер Т., Йоханссон Л., Салмонссон С.: Наблюдение за смертностью и заболеваемостью раком среди шведских рабочих с графитовыми электродами. Environ Res. 1995, 70 (1): 7-10. 10.1006 / enrs.1995.1039.
CAS Статья PubMed Google ученый
Донато Ф., Монарка С., Маркионна Дж., Росси А., Чичони С., Кьеза Р., Колин Д., Боффетта П. Смертность от рака и хронических респираторных заболеваний среди рабочих, производящих угольные электроды.Occup Environ Med. 2000, 57 (7): 484-487. 10.1136 / oem.57.7.484.
CAS Статья PubMed PubMed Central Google ученый
МАИР: Полиядерные ароматические соединения, Часть 1, Химические, экологические и экспериментальные данные. Монография МАИР по оценке канцерогенного риска химических веществ для человека. 1983, Лион (Франция): Международное агентство по изучению рака, 32:
Google ученый
Crofts F, Taioli E, Trachman J, Cosma GN, Currie D, Toniolo P, Garte SJ: Функциональное значение различных генотипов CYP1A1 человека. Канцерогенез. 1994, 15 (12): 2961-2963. 10.1093 / carcin / 15.12.2961.
CAS Статья PubMed Google ученый
Коулз Б., Кеттерер Б. Роль глутатиона и трансфераз глутатиона в химическом канцерогенезе. Crit Rev BiochemMol Biol. 1990, 25 (1): 47-70.
CAS Статья Google ученый
Ingelman-Sundberg M: Генетическая изменчивость восприимчивости и реакции на токсиканты. Toxicol Lett. 2001, 120 (1-3): 259-268. 10.1016 / S0378-4274 (01) 00278-8.
CAS Статья PubMed Google ученый
Паванелло С., Клонферо Э .: Биологические индикаторы генотоксического риска и метаболических полиморфизмов. Mutat Res. 2000, 463 (3): 285-308. 10.1016 / S1383-5742 (00) 00051-X.
CAS Статья PubMed Google ученый
Schoket B, Papp G, Levay K, Mrackova G, Kadlubar FF, Vincze I: Влияние метаболических генотипов на уровни биомаркеров генотоксического воздействия. Mutat Res. 2001, 482 (1-2): 57-69.
CAS Статья PubMed Google ученый
Bartsch H, Nair U, Risch A, Rojas M, Wikman H, Alexandrov K: Генетический полиморфизм генов CYP, отдельно или в комбинации, как модификатор риска рака, связанного с табаком. Биомаркеры эпидемиологии рака Пред.2000, 9 (1): 3-28.
CAS PubMed Google ученый
Pastorelli R, Guanci M, Cerri A, Negri E, La Vecchia C, Fumagalli F, Mezzetti M, Cappelli R, Panigalli T, Fanelli R, et al: Влияние наследственных полиморфизмов на глутатион S-трансферазу M1 , микросомальная эпоксидгидролаза, ферменты цитохрома P450 на ДНК и белковые аддукты бензо (а) пирендиолепоксида. Биомаркеры эпидемиологии рака Пред. 1998, 7 (8): 703-709.
CAS PubMed Google ученый
Рохас М., Александров К., Каскорби И., Брокмоллер Дж., Лихачев А., Пожарисский К., Бувье Г., Обуртин Г., Майер Л., Копп-Шнайдер А. и др.: Диол-эпоксид с высоким содержанием бензо [а] пирена Уровни аддуктов ДНК в легких и клетках крови людей с комбинированными генотипами CYP1A1 MspI / Msp-GSTM1 * 0 / * 0. Фармакогенетика. 1998, 8 (2): 109-118. 10.1097 / 00008571-199804000-00003.
CAS Статья PubMed Google ученый
Тейшейра Дж. П., Гаспар Дж., Мартиньо Дж., Сильва С., Родригес С., Майан О., Мартин Е., Фермер П. Б., Рюфф Дж.: Уровни аддуктов ароматической ДНК у рабочих коксовой печи: корреляция с полиморфизмами генов GSTP1, GSTM1, GSTT1 и CYP1A1. Mutat Res. 2002, 517 (1-2): 147-155.
CAS Статья PubMed Google ученый
Au WW, Oh HY, Grady J, Salama SA, Heo MY: Использование генетической предрасположенности и биомаркеров для оценки риска для здоровья, связанного с окружающей средой.Environ Mol Mutagen. 2001, 37 (3): 215-225. 10.1002 / em.1030.
CAS Статья PubMed Google ученый
Autrup H: Генетические полиморфизмы в ферментах метаболизма xenobiotica человека как факторы чувствительности при токсической реакции. Mutat Res. 2000, 464 (1): 65-76.
CAS Статья PubMed Google ученый
Раака С., Хассет Ч., Омиенсински К.Дж .: Микросомальная эпоксидгидролаза человека: генетический полиморфизм 5′-фланкирующей области.Канцерогенез. 1998, 19 (3): 387-393. 10.1093 / carcin / 19.3.387.
CAS Статья PubMed Google ученый
Борд П., Когган М., Джонстон П., Росс В., Сузуки Т., Уэбб Г.: Генетическая гетерогенность трансфераз глутатиона человека: комплекс семейств генов. Pharmacol Ther. 1990, 48 (3): 357-369. 10.1016 / 0163-7258 (90)
-6.
CAS Статья PubMed Google ученый
Бинкова Б., Топинка Дж., Мрацкова Г., Гайдосова Д., Видова П., Ставкова З., Петерка В., Пильчик Т., Римар В., Добиас Л. и др.: Исследование рабочих коксовой печи: влияние воздействия и генотипы GSTM1 и NAT2 на ДНК уровни аддукта в лейкоцитах и лимфоцитах, определяемые с помощью 32P-постмаркировки. Mutat Res. 1998, 416 (1-2): 67-84.
CAS Статья PubMed Google ученый
Alexandrie AK, Warholm M, Carstensen U, Axmon A, Hagmar L, Levin JO, Ostman C, Rannug A: Полиморфизмы CYP1A1 и GSTM1 влияют на уровни 1-гидроксипирена в моче после воздействия ПАУ.Канцерогенез. 2000, 21 (4): 669-676. 10.1093 / carcin / 21.4.669.
CAS Статья PubMed Google ученый
Срам Р.Дж., Бинкова Б. Исследования молекулярной эпидемиологии воздействия мутагенов и канцерогенов на рабочем месте и в окружающей среде, 1997–1999. Перспектива здоровья окружающей среды. 2000, 108 (Приложение 1): 57-70. 10.2307 / 3454632.
CAS Статья PubMed PubMed Central Google ученый
Jongeneelen FJ, Anzion RB, Henderson PT: Определение гидроксилированных метаболитов полициклических ароматических углеводородов в моче. J Chromatogr. 1987, 413: 227-232. 10.1016 / 0378-4347 (87) 80230-X.
CAS Статья PubMed Google ученый
VanRooij JG, Bodelier-Bade MM, Jongeneelen FJ: Оценка индивидуального кожного и респираторного поглощения полициклических ароматических углеводородов у 12 работников коксовых печей. Br J Ind Med.1993, 50 (7): 623-632.
CAS PubMed PubMed Central Google ученый
Tice RR, Agurell E, Anderson D, Burlinson B, Hartmann A, Kobayashi H, Miyamae Y, Rojas E, Ryu JC, Sasaki YF: Анализ одноклеточного геля / кометы: руководство для in vitro и in vivo генетическое токсикологическое тестирование. Environ Mol Mutagen. 2000, 35 (3): 206-221. 10.1002 / (SICI) 1098-2280 (2000) 35: 3 <206 :: AID-EM8> 3.0.CO; 2-J.
CAS Статья PubMed Google ученый
Pastorelli R, Cerri A, Rozio M, Dell’Omo M, Muzi G, Abbritti G, Fanelli R, Airoldi L: Аддукты бензо (а) пирендиолепоксида к альбумину у рабочих, подвергающихся воздействию полициклических ароматических углеводородов: связь со специфическими CYP1A1, GSTM1 , Генотипы GSTP1 и EHPX. Биомаркеры. 2001, 6 (5): 357-374. 10.1080 / 13547500110044267.
CAS Статья PubMed Google ученый
Hayashi S, Watanabe J, Kawajiri K: Генетические полиморфизмы в 5′-фланкирующей области изменяют регуляцию транскрипции гена цитохрома P450IIE1 человека.J Biochem (Токио). 1991, 110 (4): 559-565.
CAS Google ученый
Cascorbi I, Brockmoller J, Roots I: Полиморфизм C4887A в экзоне 7 человеческого CYP1A1: популяционная частота, мутации и влияние на восприимчивость к раку легких. Cancer Res. 1996, 56 (21): 4965-4969.
CAS PubMed Google ученый
Белл Д.А., Тейлор Дж. А., Полсон Д. Ф., Робертсон К. Н., Молер Дж. Л., Люсьер Г. В.: Генетический риск и воздействие канцерогенов: распространенный наследственный дефект гена метаболизма канцерогенов, глутатион-S-трансферазы M1 (GSTM1), который увеличивает предрасположенность к раку мочевого пузыря.J Natl Cancer Inst. 1993, 85 (14): 1159-1164. 10.1093 / jnci / 85.14.1159.
CAS Статья PubMed Google ученый
Brendler-Schwaab SY, Schmezer P, Liegibel U, Weber S, Michalek K, Tompa A, Pool-Zobel BL: Клетки различных тканей для токсикологических исследований in vitro и in vivo: Сборник методов выделения. Токсикология in vitro. 1994, 8 (6): 1285-1302. 10.1016 / 0887-2333 (94)
-8.
CAS Статья PubMed Google ученый
Jones KH, Senft JA: Улучшенный метод определения жизнеспособности клеток путем одновременного окрашивания флуоресцеина диацетат-пропидий йодид. J Histochem Cytochem. 1985, 33 (1): 77-79.
CAS Статья PubMed Google ученый
Сингх Н.П., Маккой М.Т., Тайс Р.Р., Шнайдер Э.Л.: простой метод количественного определения низких уровней повреждения ДНК в отдельных клетках. Exp Cell Res. 1988, 175 (1): 184-191. 10.1016 / 0014-4827 (88)-0.
CAS Статья PubMed Google ученый
Villarini M, Moretti M, Pasquini R, Scassellati-Sforzolini G, Fatigoni C, Marcarelli M, Monarca S, Rodriguez AV: Генотоксические эффекты инсектицида дельтаметрина in vitro на лейкоциты периферической крови человека: повреждение ДНК (‘ комета) в отношении индукции обменов сестринских хроматид и микроядер. Токсикология. 1998, 130 (2-3): 129-139. 10.1016 / S0300-483X (98) 00097-3.
CAS Статья PubMed Google ученый
Villarini M, Scassellati-Sforzolini G, Moretti M, Pasquini R: Генотоксичность тербутрина in vitro, оцененная с помощью щелочного одноклеточного микрогелевого электрофореза «кометного» анализа. Cell Biol Toxicol. 2000, 16 (5): 285-292. 10.1023 / А: 1026794213308.
CAS Статья PubMed Google ученый
Moretti M, Villarini M, Scassellati-Sforzolini G, Monarca S, Salucci A, Vicent Rodriguez A: Применение одноклеточного гель-электрофореза («комета») для обнаружения первичного повреждения ДНК в рабочие резиновой промышленности: сравнение ручного и компьютерного анализа.Токсикологическая и экологическая химия. 1999, 13-24.
Google ученый
Бонасси С., Фонтана В., Сеппи М., Барале Р., Бигжери А.: Анализ коррелированных данных в исследованиях биомониторинга человека. Случай клеток с высокой частотой обмена сестринских хроматид. Mutat Res. 1999, 438 (1): 13-21.
CAS Статья PubMed Google ученый
Коллинз А.Р.: Кометный анализ повреждений и репарации ДНК: принципы, применения и ограничения.Mol Biotechnol. 2004, 26 (3): 249-261. 10.1385 / МБ: 26: 3: 249.
CAS Статья PubMed Google ученый
Nan HM, Kim H, Lim HS, Choi JK, Kawamoto T., Kang JW, Lee CH, Kim YD, Kwon EH: Влияние профессии, образа жизни и генетического полиморфизма CYP1A1, CYP2E1, GSTM1 и GSTT1 на концентрации 1-гидроксипирена и 2-нафтола в моче. Канцерогенез. 2001, 22 (5): 787-793. 10.1093 / carcin / 22.5.787.
CAS Статья PubMed Google ученый
Ovrebo S, Haugen A, Fjeldstad PE, Hemminki K, Szyfter K: Биологический мониторинг воздействия полициклических ароматических углеводородов на заводе по производству электродной пасты. J Occup Med. 1994, 36 (3): 303-310. 10.1097 / 00043764-199403000-00007.
CAS Статья PubMed Google ученый
Siwinska E, Mielzynska D, Kapka L: Связь между 1-гидроксипиреном в моче и генотоксическими эффектами у работников коксовых печей. Occup Environ Med.2004, 61 (3): e10-10.1136 / oem.2002.006643.
CAS Статья PubMed PubMed Central Google ученый
Duez P, Dehon G, Kumps A, Dubois J: Статистика анализа комет: ключ к различению генотоксических эффектов. Мутагенез. 2003, 18 (2): 159-166. 10.1093 / mutage / 18.2.159.
CAS Статья PubMed Google ученый
Buschini A, De Palma G, Poli P, Martino A, Rossi C, Mozzoni P, Scotti E, Buzio L, Bergamaschi E, Mutti A: Генетический полиморфизм ферментов метаболизма лекарств и индуцированное стиролом повреждение ДНК .Environ Mol Mutagen. 2003, 41 (4): 243-252. 10.1002 / em.10150.
CAS Статья PubMed Google ученый
Лу Й, Моримото К., Накаяма К.: Практика здоровья и повреждение ДНК лейкоцитов у японских рабочих, занимающихся твердосплавными работами. Предыдущая Мед. 2006, 43 (2): 140-144. 10.1016 / j.ypmed.2006.03.008.
CAS Статья PubMed Google ученый
Sigurdson AJ, Hauptmann M, Alexander BH, Doody MM, Thomas CB, Struewing JP, Jones IM: повреждение ДНК при раке щитовидной железы и множественные случаи рака, контрольные и долгоживущие люди.Mutat Res. 2005, 586 (2): 173-188.
CAS Статья PubMed Google ученый
Carstensen U, Hou SM, Alexandrie AK, Hogstedt B, Tagesson C, Warholm M, Rannug A, Lambert B, Axmon A, Hagmar L: Влияние генетического полиморфизма ферментов биотрансформации на мутации генов, разрывы цепей дезоксирибонуклеиновая кислота и микроядра в мононуклеарных клетках крови и 8-гидроксидезоксигуанозин в моче у рабочих, подвергшихся воздействию полиароматических углеводородов.Scand J Work Environ Health. 1999, 25 (4): 351-360.
CAS Статья PubMed Google ученый
Crebelli R, Carta P, Andreoli C, Aru G, Dobrowolny G, Rossi S, Zijno A: Биомониторинг работников первичной алюминиевой промышленности: обнаружение микроядер и восстанавливаемые повреждения ДНК с помощью щелочного SCGE. Mutat Res. 2002, 516 (1-2): 63-70.
CAS Статья PubMed Google ученый
Marczynski B, Rihs HP, Rossbach B, Holzer J, Angerer J, Scherenberg M, Hoffmann G, Bruning T., Wilhelm M: анализ 8-оксо-7,8-дигидро-2′-дезоксигуанозина и разрывов цепей ДНК в белом цвете клетки крови рабочих, подвергшихся профессиональному облучению: сравнение с мониторингом окружающей среды, метаболиты в моче и полиморфизм ферментов. Канцерогенез. 2002, 23 (2): 273-281. 10.1093 / carcin / 23.2.273.
CAS Статья PubMed Google ученый
Moretti M, Villarini M, Scassellati-Sforzolini G, Monarca S, Libraro M, Fatigoni C, Donato F, Leonardis C, Perego L: Биологический мониторинг генотоксической опасности у рабочих резиновой промышленности. Перспектива здоровья окружающей среды. 1996, 104 (Приложение 3): 543-545. 10.2307 / 3432820.
CAS Статья PubMed PubMed Central Google ученый
Sul D, Oh E, Im H, Yang M, Kim CW, Lee E: Повреждение ДНК в T- и B-лимфоцитах и гранулоцитах у рабочих, занимающихся инспекцией выбросов и сжиганием, подвергшихся воздействию полициклических ароматических углеводородов.Mutat Res. 2003, 538 (1-2): 109-119.
CAS Статья PubMed Google ученый
van Delft JH, Steenwinkel MS, van Asten JG, de Vogel N, Bruijntjes-Rozier TC, Schouten T., Cramers P, Maas L, van Herwijnen MH, van Schooten F, et al: Биологический мониторинг воздействия к полициклическим ароматическим углеводородам коксоводов в связи с курением и генетическим полиморфизмом для GSTM1 и GSTT1. Ann Occup Hyg. 2001, 45 (5): 395-408.
CAS Статья PubMed Google ученый
Hoffmann H, Hogel J, Speit G: Влияние курения на эффекты ДНК в анализе комет: метаанализ. Мутагенез. 2005, 20 (6): 455-466. 10.1093 / mutage / gei064.
CAS Статья PubMed Google ученый
Cosma G, Crofts F, Currie D, Wirgin I, Toniolo P, Garte SJ: Расовые различия в полиморфизмах длины рестрикционных фрагментов и индуцируемости матричной РНК гена CYP1A1 человека.Биомаркеры эпидемиологии рака Пред. 1993, 2 (1): 53-57.
CAS PubMed Google ученый
Чжао Х., Шпиц М.Р., Гвин К.М., Ву Х: Микросомальные полиморфизмы эпоксидгидролазы и риск рака легких у неиспаноязычных белых. Mol Carcinog. 2002, 33 (2): 99-104. 10.1002 / mc.10023.
CAS Статья PubMed Google ученый
Hirvonen A, Husgafvel-Pursiainen K, Anttila S, Vainio H: нулевой генотип GSTM1 как модификатор потенциального риска плоскоклеточного рака легкого.Канцерогенез. 1993, 14 (7): 1479-1481. 10.1093 / carcin / 14.7.1479.
CAS Статья PubMed Google ученый
Somorovska M, Jahnova E, Tulinska J, Zamecnikova M, Sarmanova J, Terenova A, Vodickova L, Liskova A, Vallova B, Soucek P, et al: Биомониторинг профессионального воздействия стирола на заводе по ламинированию пластмасс . Mutat Res. 1999, 428 (1-2): 255-269.
CAS Статья PubMed Google ученый
Кульюкка-Рабб Т., Нюлунд Л., Вааранринта Р., Савела К., Мутанен П., Вейдебаум Т., Сорса М., Раннуг А., Пелтонен К.: Влияние соответствующих генотипов на биомаркеры, связанные с воздействием ПАУ. J Expo Anal Environ Epidemiol. 2002, 12 (1): 81-91. 10.1038 / sj.jea.7500204.
CAS Статья PubMed Google ученый
Апостоли П., Нери Дж., Лукас Д., Манно М., Берту Ф .: Влияние генетических полиморфизмов CYP1A1 и GSTM1 на уровни 1-гидроксипирена в моче.Toxicol Lett. 2003, 144 (1): 27-34. 10.1016 / S0378-4274 (03) 00227-3.
CAS Статья PubMed Google ученый
Pavanello S, Pulliero A, Siwinska E, Mielzynska D, Clonfero E: Пониженная эксцизионная репарация нуклеотидов и GSTM1-нулевые генотипы влияют на уровни аддукта анти-B [a] PDE-ДНК в мононуклеарных белых кровяных тельцах с высоким содержанием PAH -открытые работники коксовых печей. Канцерогенез. 2005, 26 (1): 169-175. 10.1093 / carcin / bgh403.
CAS Статья PubMed Google ученый
Рохас М., Каскорби И., Александров К., Крик Э., Обуртин Г., Майер Л., Копп-Шнайдер А., Рутс I, Барч Н.: Модуляция уровней аддукта бензо [а] пирендиолепоксида-ДНК в лейкоцитах человека с помощью CYP1A1, Полиморфизм GSTM1 и GSTT1. Канцерогенез. 2000, 21 (1): 35-41. 10.1093 / carcin / 21.1.35.
CAS Статья PubMed Google ученый
Cotelle S, Ferard JF: Анализ комет в генетической экотоксикологии: обзор. Environ Mol Mutagen.1999, 34 (4): 246-255. 10.1002 / (SICI) 1098-2280 (1999) 34: 4 <246 :: AID-EM4> 3.0.CO; 2-V.
CAS Статья PubMed Google ученый
Merlo DF, Garattini S, Gelatti U, Simonati C, Covolo L, Ceppi M, Donato F: когортное исследование смертности среди рабочих на заводе по производству графитовых электродов в Италии. Occup Environ Med. 2004, 61 (2): e9-10.1136 / oem.2003.009357.
CAS Статья PubMed PubMed Central Google ученый
| Штамм | Идентификационный номер модели гена | Тип элемента | Координаты | Отборные штаммы |
|---|---|---|---|---|
| C57BL / 6J | MGI_C57BL6J_95405 | ген, кодирующий белок | Chr1: 180803775-180848469 (-) | |
| 129S1 / SvImJ | MGP_129S1SvImJ_G0016880 | ген, кодирующий белок | Chr1: 187460320-187506279 (-) | |
| A / J | MGP_AJ_G0016859 | ген, кодирующий белок | Chr1: 179869402-1798 (-) | |
| AKR / J | MGP_AKRJ_G0016825 | ген, кодирующий белок | Chr1: 185215050-185261915 (-) | |
| BALB / cJ | MGP_BALBcJ_G0016817 | ген, кодирующий белок | Chr1: 1804-180534911 (-) | |
| C3H / HeJ | MGP_C3HHeJ_G0016642 | ген, кодирующий белок | Chr1: 185989296-186035480 (-) | |
| C57BL / 6NJ | MGP_C57BL6NJ_G0017280 | ген, кодирующий белок | Chr1: 193548432-193600390 (-) | |
| КАРОЛИ / ЭЙДЖ | MGP_CAROLIEiJ_G0014988 | ген, кодирующий белок | Chr1: 170842563-170887301 (-) | |
| CAST / EiJ | MGP_CASTEiJ_G0016223 | ген, кодирующий белок | Chr1: 185334546-185383637 (-) | |
| CBA / J | MGP_CBAJ_G0016616 | ген, кодирующий белок | Chr1: 200680779-200728941 (-) | |
| DBA / 2J | MGP_DBA2J_G0016722 | ген, кодирующий белок | Chr1: 178702879-178747320 (-) | |
| FVB / NJ | MGP_FVBNJ_G0016718 | ген, кодирующий белок | Chr1: 176802928-176847816 (-) | |
| LP / J | MGP_LPJ_G0016796 | ген, кодирующий белок | Chr1: 188710360-188761275 (-) | |
| NOD / ShiLtJ | MGP_NODShiLtJ_G0016749 | ген, кодирующий белок | Chr1: 207848003-207893903 (-) | |
| NZO / HlLtJ | MGP_NZOHlLtJ_G0017317 | ген, кодирующий белок | Chr1: 184993673-185042577 (-) | |
| PWK / PhJ | MGP_PWKPhJ_G0016011 | ген, кодирующий белок | Chr1: 177819463-177864661 (-) | |
| SPRET / EiJ | MGP_SPRETEiJ_G0015787 | ген, кодирующий белок | Chr1: 183929976-183989268 (-) | |
| WSB / EiJ | MGP_WSBEiJ_G0016286 | ген, кодирующий белок | Chr1: 185196660-185243798 (-) |
Частоты генетических полиморфизмов в гене EPHX в контроле…
Contexte. Au Cours de la pre-éclampsie (PE), le défaut d’invasion trophoblastique et de remodelage des artères utérines spiralées génère une mauvaise, адаптация де-ла циркуляции утеро-плаценты, ассоциативно связанной с phénomènes d’ypoxie / réoxygén. Оксидант стресса и нарушение равновесия ангиогенных / антиангиогенных факторов (уменьшение VEGF и PIGF по сравнению с увеличением sFlt1) отвечает за нормальную плацентацию, за нарушение эндотелиальной функции и за нарушение системы воспаления.Дисфункция эндотелиального оксида нитриксинтазы (eNOS) и уменьшение биодисперсности NO, является критикой роли патофизиологии ПЭ. eNOS — это основной источник продукции NO в плаценте, а также в изначальной роли в гомеостазии и регуляции тонуса сосудов. Des données récentes indiquent que eNOS peut être modifiée au Cours du стресс-оксидант, особенно в отношении S-глутатионилирования, который вступает в реакцию с последующим разобщением с образованием супероксида и восстановлением NO.Objectif. Le but de ce travail a été d’étudier les conséquences du стресс-оксидант на eNOS-плацентар, в частности, на глутатионилирование, и другие модификации, связанные с продуктом липидного окисления (LPO), в отношении avec sa disfonction observée au Cours de la PE . Matériels et Méthodes. Модификация eNOS a été étudiée dans des Tissus Placentaires obtenus à partir de césariennes réalisées chez des Patientes pre-éclamptiques (n = 13), vs grosses normales (n = 9). Дополнительные материалы, созданные для людей с трофобластами, HTR-8 / SVneo обнажают оксидант стресса, индуцирующий H / R или показывающий LPO.Результаты: Исследования по иммунофлуоресценции и микроскопии, конфокальной, наблюдающей за важным глутатионилированием eNOS в плаценте PE, разумными вспомогательными веществами-возбудителями (дитиотреитол), без различия в выражении eNOS в целом в PE и в больших количествах. S-глутатионилирование подтверждается иммунопреципитацией и вестерн-блоттингом плаценты eNOS. Экспозиция трофобластов HTR8 в условиях H / R, связанная с S-глутатионилированием NO, связана с образованием NO и образованием супероксида.Le NO необходим для потенциального инвазии трофобластов, при отсутствии миграции HTR8 после специального ингибирования eNOS в отношении малых ARN-интерференций (siRNA) и после добавления NO, NOC-18. Les trophoblastes демонстрирует дополнительные вариации H / R, текущие способности к уменьшению миграции, временные инвазивные и возобновляемые NOC-18. Dans la deuxième partie de ce travail, nous avons recherché la présence de LPO dans les placentas de PE, et nous avons émis l’hypothèse que eNOS puisse être un cible de ces агентов.Nous montrons que les LPO tels que le 4-hydroxynonenal (4-HNE), et le 4-oxo-2-nonenal (ONE), s’accumulent dans les placentas de PE, особенно на eNOS, alors qu’aucune модификации N’est Observée dans les placentas de grossesse normale. Исследования в области защиты рекомбинантных eNOS, текущих ONE и 4-HNE-модификаторов с дополнительными эпитопами (ONE-Lys, HNE-His, HNE-Cys). Добавление 4-HNE или ONE aux HTR8, ингибирующее производство NO и миграцию клеток, восстанавливается вместе с добавлением NOC-18.