LADA Vesta SW – Технические характеристики – Официальный сайт LADA
Колесная формула / ведущие колеса
Расположение двигателя
Тип кузова / количество дверей
Количество мест
Длина / ширина / высота по антенне, мм
База, мм
Колея передних / задних колес, мм
Дорожный просвет, мм

Объем багажного отделения в пассажирском / грузовом…
Код двигателя
Тип двигателя
Система питания
Количество, расположение цилиндров
Рабочий объем, куб. см
Максимальная мощность, кВт (л.с.) / об. мин.
Максимальный крутящий момент, Нм / об. мин.
Рекомендуемое топливо
Максимальная скорость, км/ч
Время разгона 0-100 км/ч, с
Городской цикл, л/100 км
Загородный цикл, л/100 км
Смешанный цикл, л/100 км
Снаряженная масса, кг
Технически допустимая максимальная масса, кг
 ..»>
..»>Максимальная масса прицепа без тормозной системы /…
Объем топливного бака, л
Тип трансмиссии
Передаточное число главной передачи
Передняя
Задняя
Рулевой механизм
Размерность
Какой объем багажника на ВАЗ-2114
ВАЗ-2114 – машина, которая пришла на замену «девятки», снятой с производства, и сразу завоевала популярность и стала востребованной. По сравнению со своей предшественницей, данная модель автомобиля имеет несколько отличий. Различия коснулись не только внешних частей кузова, но также салона. В частности, у ВАЗ-2114 большой объем багажника. Накладки на порогах, молдинги вдоль бортов, спойлер со стоп-сигналом – все это придает машине современный внешний вид.
Различия коснулись не только внешних частей кузова, но также салона. В частности, у ВАЗ-2114 большой объем багажника. Накладки на порогах, молдинги вдоль бортов, спойлер со стоп-сигналом – все это придает машине современный внешний вид.
Внешние изменения отразились на эксплуатационных характеристиках машины. В частности, данная модель обладает прекрасными аэродинамическими качествами. Она устойчива на дороге, даже если движение осуществляется на высокой скорости. Сочетание цветовой гаммы кузова и навесных деталей придает этому автомобилю вид иномарки.
Благодаря внешнему виду эта машина вызвала повышенный интерес у людей, которые занимаются тюнингом. При сравнительно небольших финансовых затратах из такого автомобиля можно запросто сделать выделяющуюся из общей массы оригинальную машину, подчеркнув тем самым индивидуальность и чувство стиля ее владельца.
Багажник
За счет возможности, при которой можно сложить заднее сидение, этот автомобиль легко трансформируется в грузопассажирское транспортное средство. У данной модели авто объем багажника равен:
У данной модели авто объем багажника равен:
- 330 дм 3 при разложенных сиденьях;
- 600 дм 3 при сложенных задних сиденьях.
В литрах объем такой же, как в кубах (кубических дециметрах).
Такая вместительность машины позволяет перевозить в ней даже габаритные грузы, что особенно актуально для тех, кто часто сталкивается с необходимостью доставить какой-либо товар или груз другого назначения до места выгрузки. При сложенных сиденьях объем багажника увеличивается почти в 2 раза.
Кроме того, если требуется перевезти большое количество груза, или, например, вы отправляетесь в поездку со всей семьей, вам не придется ограничиваться местом объемом 330 л для расположения вещей. В продаже доступны различные крепления и установки для этой модели автомобиля, которые позволяют перевозить куда больше вещей, чем поместится в багажное отделение.
Например, вы можете установить багажник на крышу. В продаже доступны различные варианты таких креплений:
- Автобоксы.

- Крепления для спортивного снаряжения.
- Корзины.
- Экспедиционные багажники на крышу.

Таким образом, вам удастся сделать свой автомобиль еще более вместительным и освободить место для пассажиров.
Управление с кнопки
Очень удобно, когда управление (открытие багажника) осуществляется с кнопки. Каждый автовладелец знает, что очень важно позаботиться о том, чтобы багажник, как и двери автомобиля, надежно закрывались. Надежная работа замков багажника важна так же, как и надежность замка в двери гаража или квартиры. Все современные модели машин оснащены механизмом открытия багажника с помощью пульта управления или специальной кнопки. Установка механизма с кнопкой не просто делает управление более удобным, но и решает ряд проблем. В частности, таким образом решается проблема обледенения замка, который открывается ключом.
Установку такого механизма можно доверить специалисту. Но некоторые автовладельцы предпочитают выполнять эту работу самостоятельно. Для этого понадобится стальной трос, ролик, кнопка подключения, электромагнит привода багажника и провода. Ролик устанавливается на корпусе редуктора стеклоочистителя. Если ролик открытый, вы можете использовать трос в оплетке. Самый простой способ подключения – к прикуривателю. Для других вариантов требуется иметь соответствующие знания электросхемы.
Для этого понадобится стальной трос, ролик, кнопка подключения, электромагнит привода багажника и провода. Ролик устанавливается на корпусе редуктора стеклоочистителя. Если ролик открытый, вы можете использовать трос в оплетке. Самый простой способ подключения – к прикуривателю. Для других вариантов требуется иметь соответствующие знания электросхемы.
Автомобили ВАЗ, 2113, 2114 и 2115 – модели, пользующиеся популярностью у отечественных водителей. Это связано с их невысокой стоимостью, ремонтопригодностью, относительной надежностью и неприхотливостью. А от объема багажника ВАЗ 2114 зависит практичность эксплуатации машины.
ВАЗ 2113
Благодаря объему багажника ВАЗ 2113, у водителей есть возможность перевозить габаритные грузы. Его вместительность – 330 литров. Технические характеристики багажного отделения предоставляют возможность перевозить предметы, имеющие нестандартные размеры.
Багажник модели 2113 трансформируется: водитель может убрать полку, сложить задние сиденья. После этого его вместимость составляет 632 литра. Хэтчбек позволяет перевозить малогабаритные, длинномерные грузы.
К недостаткам относят отсутствие штатной подсветки (при тюнинге авто ее монтируют самостоятельно), высокий погрузочный бортик, который осложняет помещение грузов вовнутрь.
ВАЗ 2114
ВАЗ 2114 – надежный пятидверный автомобиль. Вместимость составляет 330 литров при сложенных сиденьях, а при разложенных – она равна 632 литрам. Погрузочная высота бортика имеет неудобный уровень (аналогично тому, который используется на тринадцатой модели), что существенно усложняет процесс погрузки.
Габариты багажника выглядят следующим образом: длина – 830 миллиметров при сложенных сиденьях и практически 1660 миллиметров – при разложенных, ширина – 935 миллиметров, глубина – 830 миллиметров, высота – 780 миллиметров. Эти размеры приблизительные и актуальны для моделей 2113 и 2114.

Возможности трансформации позволяют загружать предметы, чьи длина, а также размеры отличаются от стандартных габаритов. При установке дополнительных багажных конструкций на крышу можно перевезти определенное количество крупногабаритных грузов.
Вместимость, выраженная в литрах, идентична объему, который указывается в кубических дециметрах.
ВАЗ 2115
Объем багажника ВАЗ 2115 составляет 427 литров при сложенных сиденьях, если их разложить, то вместимость увеличивается до 700 литров. Габариты багажного отделения увеличены, благодаря чему даже крупногабаритные грузы могут быть перевезены на пятнадцатой модели.
К преимуществам относят простоту демонтажа, установку полки, а также удобную высоту для погрузки. Она находится практически на уровне автомобильного бампера.
Часто водители улучшают шумоизоляцию и штатную обшивку багажного отделения. Монтируют электрический привод открывания крышки, устанавливают подсветку, конструируют боксы для хранения небольших предметов (однако за счет их установки, багажник существенно теряет объем).

Особенности багажника
К особенностям багажных отделений различных модификаций моделей 2113, 2114, 2115 относят приемлемый уровень вместимости и возможность трансформации салона для увеличения полезного объема.
ВАЗ 2115 отличает удобная погрузочная высота, более увеличенный объем багажника Лада 2115. В остальном конструкция отделения для перевозки груза идентична другим моделям.
Авто оснащаются штатной полкой, она устанавливается вертикально и разделяет багажное отделение и салон. В случае необходимости (например, при перевозке крупногабаритных грузов, требующих складывания задних сидений), эту полку и вертикальную шторку можно демонтировать.
На автомобилях Лада 2113, 2114, 2115 под «полом» багажника предусмотрена ниша для хранения запасного колеса и инструментов, необходимых для быстрой починки небольших неисправностей, которые могут возникнуть в пути.
ВАЗ 2115 отличает удобная погрузочная высота, более увеличенный объем багажника Лада 2115. В остальном его конструкция идентична другим моделям.
В остальном его конструкция идентична другим моделям.
Модели ВАЗ 2113, 2114, 2115, являющиеся модификацией «восьмерок», «девяток» и «девяносто девятых», соответственно, стали более эргономичными, удобными в использовании, практичными. Автомобили получили просторные багажные отделения, благодаря чему увеличилось количество перевозимого груза. Наличие ряда недостатков не отпугивает водителей: некоторые неудобства можно исправить самостоятельно.
Автомобили семейства ВАЗ 2113, 2114, 2115 популярны не только в России, но и в государствах СНГ. Это связано с большим распространением этих авто, их неприхотливостью, относительной надежностью, ремонтопригодностью и стоимостью. Машины обладают приемлемой вместимостью погрузочной ниши, что делает их удобными в использовании для решения повседневных бытовых задач или ведения небольшого бизнеса.
Автомобили ВАЗ, 2113, 2114 и 2115 – модели, пользующиеся популярностью у отечественных водителей. Это связано с их невысокой стоимостью, ремонтопригодностью, относительной надежностью и неприхотливостью. А от объема багажника ВАЗ 2114 зависит практичность эксплуатации машины.
А от объема багажника ВАЗ 2114 зависит практичность эксплуатации машины.
Багажник модели 2113 трансформируется: водитель может убрать полку, сложить задние сиденья. После этого его вместимость составляет 632 литра. Хэтчбек позволяет перевозить малогабаритные, длинномерные грузы.
К недостаткам относят отсутствие штатной подсветки (при тюнинге авто ее монтируют самостоятельно), высокий погрузочный бортик, который осложняет помещение грузов вовнутрь.
ВАЗ 2114
ВАЗ 2114 – надежный пятидверный автомобиль. Вместимость составляет 330 литров при сложенных сиденьях, а при разложенных – она равна 632 литрам. Погрузочная высота бортика имеет неудобный уровень (аналогично тому, который используется на тринадцатой модели), что существенно усложняет процесс погрузки.
Габариты багажника выглядят следующим образом: длина – 830 миллиметров при сложенных сиденьях и практически 1660 миллиметров – при разложенных, ширина – 935 миллиметров, глубина – 830 миллиметров, высота – 780 миллиметров.
 Эти размеры приблизительные и актуальны для моделей 2113 и 2114.
Эти размеры приблизительные и актуальны для моделей 2113 и 2114.Возможности трансформации позволяют загружать предметы, чьи длина, а также размеры отличаются от стандартных габаритов. При установке дополнительных багажных конструкций на крышу можно перевезти определенное количество крупногабаритных грузов.
Вместимость, выраженная в литрах, идентична объему, который указывается в кубических дециметрах.
ВАЗ 2115
Объем багажника ВАЗ 2115 составляет 427 литров при сложенных сиденьях, если их разложить, то вместимость увеличивается до 700 литров. Габариты багажного отделения увеличены, благодаря чему даже крупногабаритные грузы могут быть перевезены на пятнадцатой модели.
Читайте также: Объем и размеры багажника внедорожника Рено Дастера
К преимуществам относят простоту демонтажа, установку полки, а также удобную высоту для погрузки. Она находится практически на уровне автомобильного бампера.
Она находится практически на уровне автомобильного бампера.
Часто водители улучшают шумоизоляцию и штатную обшивку багажного отделения. Монтируют электрический привод открывания крышки, устанавливают подсветку, конструируют боксы для хранения небольших предметов (однако за счет их установки, багажник существенно теряет объем).
Особенности багажника
К особенностям багажных отделений различных модификаций моделей 2113, 2114, 2115 относят приемлемый уровень вместимости и возможность трансформации салона для увеличения полезного объема.
ВАЗ 2115 отличает удобная погрузочная высота, более увеличенный объем багажника Лада 2115. В остальном конструкция отделения для перевозки груза идентична другим моделям.
Авто оснащаются штатной полкой, она устанавливается вертикально и разделяет багажное отделение и салон. В случае необходимости (например, при перевозке крупногабаритных грузов, требующих складывания задних сидений), эту полку и вертикальную шторку можно демонтировать.
На автомобилях Лада 2113, 2114, 2115 под «полом» багажника предусмотрена ниша для хранения запасного колеса и инструментов, необходимых для быстрой починки небольших неисправностей, которые могут возникнуть в пути.
ВАЗ 2115 отличает удобная погрузочная высота, более увеличенный объем багажника Лада 2115. В остальном его конструкция идентична другим моделям.
Модели ВАЗ 2113, 2114, 2115, являющиеся модификацией «восьмерок», «девяток» и «девяносто девятых», соответственно, стали более эргономичными, удобными в использовании, практичными. Автомобили получили просторные багажные отделения, благодаря чему увеличилось количество перевозимого груза. Наличие ряда недостатков не отпугивает водителей: некоторые неудобства можно исправить самостоятельно.
Автомобили семейства ВАЗ 2113, 2114, 2115 популярны не только в России, но и в государствах СНГ. Это связано с большим распространением этих авто, их неприхотливостью, относительной надежностью, ремонтопригодностью и стоимостью. Машины обладают приемлемой вместимостью погрузочной ниши, что делает их удобными в использовании для решения повседневных бытовых задач или ведения небольшого бизнеса.
Машины обладают приемлемой вместимостью погрузочной ниши, что делает их удобными в использовании для решения повседневных бытовых задач или ведения небольшого бизнеса.
Эксплуатационные характеристики ВАЗ 2114 четырнадцатая хэтчбек
Максимальная скорость: 165 км/ч
Время разгона до 100 км/ч: 13 c
Расход топлива на 100км по городу: 8.9 л
Объем бензобака: 43 л
Снаряженная масса автомобиля: 970 кг
Размер шин: 165/70 R13
Характеристики двигателя
Расположение: спереди, поперечно
Объем двигателя: 1499 см3
Мощность двигателя: 79 л.с.
Количество оборотов: 4800
Крутящий момент: 118/2800 н*м
Система питания: Распределенный впрыск
Турбонаддув: нет
Газораспределительный механизм: OHC
Расположение цилиндров: Рядный
Количество цилиндров: 4
Количество клапанов на цилиндр: 2
Рекомендуемое топливо: АИ-95
Тормозная система
Передние тормоза: Дисковые
Задние тормоза: Барабанные
Рулевое управление
Тип рулевого управления: Шестерня-рейка
Усилитель руля: нет
Трансмиссия
Привод: Передний
Количество передач: механическая коробка — 5
Передаточное отношение главной пары: 3. 7
7
Подвеска
Передняя подвеска: Амортизационная стойка
Задняя подвеска: Продольный рычаг
Кузов
Тип кузова: хэтчбек
Количество дверей: 5
Количество мест: 5
Длина машины: 4100 мм
Ширина машины: 1620 мм
Высота машины: 1415 мм
Колесная база: 2460 мм
Колея передняя: 1400 мм
Колея задняя: 1370 мм
Дорожный просвет (клиренс): 165 мм
Объем багажника: 427 л
Производство
Год выпуска: с 2001 по 2013
Lada Vesta SW Cross 2021 обзор, комплектации и цены, характеристики
Обзор LADA Vesta SW Cross
LADA Vesta SW Сross.
Скажи жизни да!
Больше возможностей на дороге и вне ее пределов – вот философия LADA Vesta SW Cross!Контрастный спортивный стиль сочетается с комфортом и отличной проходимостью: у LADA Vesta SW Cross дорожный просвет, достойный кроссовера. 203 миллиметра клиренса позволяют не переживать о высоте бордюра, глубине снега или грунтовой колеи.
203 миллиметра клиренса позволяют не переживать о высоте бордюра, глубине снега или грунтовой колеи.
Позволить себе чуть больше
LADA Vesta
Совершенство – в графике и характере
Стремительный автомобиль на мощных колесах. Высоко приподнятый кузов, окаймленный полосой защитного пластика. Пластиковый обвес порогов, арок колес и дверей защитит эмаль кузова при поездках по лёгкому бездорожью. А еще эти стильные элементы делают автомобиль зрительно крупнее!
Самый просторный в классе салон
Самый просторный в классе салон – это возможность разместиться с максимальным удобством. А полный комплект опций для достижения наибольшего комфорта позволяет сконцентрироваться на главном — удовольствии от вождения. Особое внимание уделено пассажирам – для них предусмотрен обогрев заднего сиденья, отдельный плафон освещения и USB-гнездо для подзарядки гаджетов.
Мультимедийная система
Мультимедийная система с 7-дюймовым экраном – это любимая музыка, новости по радио, телефонная связь hands free. Все, что нужно, чтобы поездка проходила быстрей и приятней! А навигатор сделает путешествие не только быстрее, но и проще, эффективней. На экран мультимедиа выводится изображение с камеры заднего вида – вместе с парктроником это обеспечивает уверенное маневрирование в тесноте городов.
Все, что нужно, чтобы поездка проходила быстрей и приятней! А навигатор сделает путешествие не только быстрее, но и проще, эффективней. На экран мультимедиа выводится изображение с камеры заднего вида – вместе с парктроником это обеспечивает уверенное маневрирование в тесноте городов.
Трансформация багажного отсека
«Органайзер на колесах» — лучший комплимент для автомобиля такого класса. Двойной пол багажника, множество ниш и крепежа для самого разнообразного груза, раскладная спинка сиденья – каждая деталь LADA Vesta SW Cross говорит о практичности семейного автомобиля. Объем багажного отделения – 480 л (до полки-шторки), а при сложенных сиденьях второго ряда – 825 л (до подоконной линии).
Высокий уровень защиты
Высокий уровень безопасности, начиная с базовой комплектации – это часть философии LADA Vesta. Система контроля устойчивости ESC позволяет уверенно маневрировать на дорогах с любым покрытием. Система экстренного оповещения ЭРА-ГЛОНАСС, мощный каркас кузова с применением сталей повышенной прочности и фронтальные подушки безопасности – это надежная защита водителя и пассажиров.
Двигатели
LADA Vesta – это возможность выбрать силовой агрегат в соответствии с индивидуальным стилем вождения и потребностями в динамике. 1,6л + «механика» — экономичный вариант. 1,8л + «механика» — солидный запас тяги! 1,6л + автоматическая трансмиссия – плавное движение и уверенный разгон! Ну а специальные настройки шасси обеспечивают комфорт и азартную управляемость.
Рабочий объем 1596 см
Мощность 106 л.с. (78 кВт) при 5800 об/мин
Момент крутящий 148 Нм при 4200 об/мин
Нормы токсичности ЕВРО-5
Рабочий объем 1774 см
Мощность 122 л.с. (90 кВт) при 5900 об/мин
Момент крутящий 170 Нм при 3700 об/мин
Нормы токсичности ЕВРО-5
Рабочий объем 1598 см
Мощность 113 л.с. (83 кВт) при 5500 об/мин
Момент крутящий 152 Нм при 4000 об/мин
Нормы токсичности ЕВРО-5
Любая модель LADA Vesta теперь может стать еще более яркой, контрастной и стильной. Достаточно заказать опционное покрытие крыши и наружных зеркал в черном глянцевом цвете. Какое сочетание смотрится лучше? Черный и красный? Черный и синий? А может, черный и оранжевый? Выбирать вам! Черная крыша применяется совместно со всей палитрой эмалей LADA Vesta, исключая «Маэстро» и «Фантом».
Какое сочетание смотрится лучше? Черный и красный? Черный и синий? А может, черный и оранжевый? Выбирать вам! Черная крыша применяется совместно со всей палитрой эмалей LADA Vesta, исключая «Маэстро» и «Фантом».
Комплектации и цены
| Подушка безопасности водителя | |||
| Подушка безопасности переднего пассажира с функцией отключения | |||
| Боковые подушки безопасности | |||
| Подголовники задних сидений 3 шт. | |||
| Крепления для детских сидений ISOFIX | |||
| Блокировка задних дверей от открывания детьми | |||
| Автоматическое запирание дверей при начале движения | |||
| Автоматическое включение аварийной сигнализации при экстренном торможении | |||
| Автоматическое отпирание дверей и включение аварийной сигнализации при столкновении | |||
| Иммобилайзер | |||
| Охранная сигнализация | |||
| Дневные ходовые огни | |||
| Противотуманные фары | |||
| Система экстренного оповещения ЭрА-ГЛОНАСС | |||
| Дисковые тормоза задних колес | |||
| Антиблокировочная система тормозов с системой вспомогательного торможения (ABS+BAS) | |||
| Электронная система распределения тормозных сил (EBD) | |||
| Система электронного контроля устойчивости (ESC) | |||
| Противобуксовочная система (TCS) | |||
| Система помощи при трогании на подъеме ^А) | |||
| Защита двигателя и подкапотного пространства | |||
| Бортовой компьютер | |||
| Подсказчик переключения передач в комбинации приборов | |||
| Центральный подлокотник с боксом | |||
| Заднее сиденье с раскладной спинкой в пропорции 60/40 | |||
| Интерьер (по выбору) Orange оранжевый/Gray серый | |||
| Противосолнечный козырек водителя и пассажира с зеркалом | |||
| Футляр для очков | |||
| Розетка 12V | |||
| Розетка 12V в багажном отделении | |||
| Двойной пол багажного отделения | |||
| Подсветка мест входа-выхода в передних дверях | |||
| Задний подлокотник | |||
| LED-подсветка интерьера | |||
| Электроусилитель рулевого управления | |||
| Регулируемая по высоте и по вылету рулевая колонка | |||
| Регулировка ремней безопасности передних сидений по высоте | |||
| Сиденье водителя с регулировкой по высоте и поясничной поддержкой | |||
| Воздушный фильтр салона | |||
| Легкая тонировка стекол | |||
| Складной ключ | |||
| Центральный замок с дистанционным управлением | |||
| Электростеклоподъемники передних дверей | |||
| Электростеклоподъемники задних дверей | |||
| Подогрев передних сидений 3х уровневый | |||
| Электропривод и электрообогрев наружных зеркал | |||
| Обогрев ветрового стекла | |||
| Датчики парковки задние | |||
| Датчики дождя и света | |||
| Климат-контроль | |||
| Охлаждаемый вещевой ящик | |||
| Круиз-контроль и ограничитель скорости | |||
| Мультифункциональное рулевое колесо | |||
| Аудиосистема (4,3» монохромный дисплей, FM/AM с функцией RDS, USB, SD-карта, AUX, Bluetooth, Hands free), 4 динамика | |||
| Камера заднего вида | |||
| Мультимедийная система с навигацией (7» цветной дисплей с TouchScreen, FM/AM с функцией RDS, USB, SD-карта, AUX, Bluetooth, Hands free), 6 динамиков | |||
| Усиленная тонировка задних стекол | |||
| Подогрев задних сидений | |||
| Наружные зеркала с боковыми указателями поворота | |||
| Наружные зеркала и наружные ручки дверей в цвет кузова | |||
| Рейлинги | |||
| Спойлер | |||
| Декоративная насадка выпускной трубы | |||
| 17» легкосплавные диски | |||
| Запасное стальное колесо временного использования 15» | |||
Характеристики
| Расположение двигателя | |||
| Длина / ширина / высота, мм | |||
| База, мм | |||
| Дорожный просвет, мм | |||
Объем багажного отделения в пассажирском / грузовом. .. .. | |||
| Колесная формула / ведущие… | |||
| Тип кузова / количество… | |||
| Количество мест | |||
| Колея передних / задних колес,… | |||
| Тип двигателя | |||
| Система питания | |||
| Рабочий объем, куб. см | |||
| Топливо | |||
| Код двигателя | |||
| Количество, расположение… | |||
| Максимальная мощность, кВт (л.с.) / об…. | |||
| Максимальный крутящий момент, Нм / об…. | |||
| Максимальная скорость, км/ч | |||
| Время разгона 0-100 км/ч, с | |||
| Городской цикл, л/100 км | |||
| Загородный цикл, л/100 км | |||
| Смешанный цикл, л/100 км | |||
| Снаряженная масса, кг | |||
| Объем топливного бака, л | |||
Технически допустимая максимальная масса,. .. .. | |||
| Максимальная масса прицепа без тормозной системы /… | |||
| Тип трансмиссии | |||
| Передаточное число главной… | |||
| Передняя | |||
| Задняя | |||
| Рулевой механизм | |||
Фотографии
Экстерьер
Интерьер
Руководства по эксплуатации
Руководство по эксплуатации LADA Vesta от 29.10.20
Руководство по эксплуатации LADA Vesta от 27.08.20
Руководство по эксплуатации LADA Vesta от 07.07.20
Руководство по эксплуатации LADA Vesta от 25.03.20
Руководство по эксплуатации LADA Vesta от 26.02.20
Дополнение к руководству по эксплуатации LADA Vesta CNG от 26. 02.20
02.20
Руководство по эксплуатации LADA Vesta от 31.01.20
Руководство по эксплуатации LADA Vesta от 15.11.19
Дополнение к руководству по эксплуатации LADA Vesta CNG от 04.06.19
Мультимедийная система LADA Vesta от 04.06.19
Руководство по эксплуатации LADA Vesta от 04.06.19
Руководство по эксплуатации LADA Vesta от 28.02.19
Руководство по эксплуатации LADA Vesta от 11.01.19
Руководство по эксплуатации LADA Vesta от 21.11.18
Дополнение к руководству по эксплуатации LADA Vesta Sport от 01.10.18
Руководство по эксплуатации LADA Vesta от 31.08.18
Руководство по эксплуатации LADA Vesta от 20.06.18
Руководство по эксплуатации LADA Vesta от 06.02.18
Руководство по эксплуатации LADA Vesta от 21.12.17
Руководство по эксплуатации LADA Vesta от 04.10.17
Руководство по эксплуатации LADA Vesta от 21. 08.17
08.17
Дополнение к руководству по эксплуатации LADA Vesta CNG от 23.06.17
Руководство по эксплуатации LADA Vesta от 05.06.17
Мультимедийная система LADA Vesta от 31.05.17
Руководство по эксплуатации LADA Vesta от 28.04.17
Руководство по эксплуатации LADA Vesta от 17.03.17
Руководство по эксплуатации LADA Vesta от 01.03.17
Мультимедийная система LADA Vesta от 13.01.17
Руководство по эксплуатации LADA Vesta от 23.12.16
Мультимедийная система LADA Vesta от 20.10.16
Руководство по эксплуатации LADA Vesta от 09.09.16
Руководство по эксплуатации LADA Vesta от 14.07.16
Руководство по эксплуатации LADA Vesta от 21.06.16
Руководство по эксплуатации LADA Vesta от 20.04.16
Руководство по эксплуатации LADA Vesta от 25.02.16
Мультимедийная система LADA Vesta от 01. 02.16
02.16
Руководство по эксплуатации LADA Vesta от 29.01.16
Руководство по эксплуатации LADA Vesta от 24.11.15
Мультимедийная система LADA Vesta от 04.11.15
| Рабочий объем, л | 1.6 | |||
| Рабочий объем, см3 | 1596 | 1597 | 1596 | |
| Диаметр цилиндра | 82 | |||
| Количество клапанов | 8 | 16 | ||
| Количество цилиндров | 4 | |||
| Максимальная мощность, кВт | 64 | 72 | 78 | |
Максимальная мощность, л. с. с. | 87 | 98 | 106 | |
| Номинальный крутящий момент, Н•м | 145 | 140 | 145 | |
| Об/мин КВТ | 0 | 5100 | 5600 | |
| Об/мин ЛС | 5800 | 5100 | 5600 | |
| Об/мин НМ | 4000 | 3800 | 4000 | |
| Расположение двигателя | переднее, поперечное | |||
| Расположение распределительного вала | DOHC | |||
| Расположение цилиндров | в ряд | |||
| Степень сжатия | 9. 8 8 | 10.5 | 10.1 | |
| Тип топлива | Бензиновый | |||
| Требования к топливу | АИ-95 | |||
| Ход поршня | 75.6 | |||
| Тип наддува | Нет | |||
| Экологический класс | EURO5 | |||
| Передняя подвеска | Независимая, «Мак-Ферсон», винтовые пружины, со стабилизатором поперечной устойчивости | |||
| Задняя подвеска | Торсионная балка, винтовые пружины | |||
| Передний амортизатор | Гидравлические | |||
| Задний амортизатор | Гидравлические | |||
Раскрыты последние секреты нового кроссовера Kia Seltos — Российская газета
Новый кроссовер Kia Seltos, несмотря на то, что был официально представлен, хранил некоторые интересные секреты: в частности, компания не раскрыла габаритов премиальной альтернативы Hyundai Creta и умолчала о характеристиках моторов для некоторых рынков. Индийскому подразделению Autocar удалось раскрыть эти секреты.
Индийскому подразделению Autocar удалось раскрыть эти секреты.
Длина Kia Seltos равна 4 315 мм, ширина — 1 800 мм, высота — 1 620 мм, колёсная база — 2 610 мм. Как и предполагалось, Seltos крупнее Hyundai Creta, чьи габариты соответственно составляют 4 270 х 1 780 х 1 665 мм, размер колёсной базы — 2 590 мм. Клиренс у моделей одинаковый — 190 мм.
Также был рассекречен объём багажника новинки — 433 литра, у Creta он почти на 30 литров меньше.
В Индии Kia Seltos будет предложен с 1.5-литровым бензиновым двигателем мощностью 115 л.с. (144 Нм), дизелем CRDi аналогичной мощности и объёма (момент, естественно, больше — 250 Нм) и турбированным 1.4 T-GDI, отдача которого составляет 140 л.с. (242 Нм).
Все они могут быть заказаны в паре с 5-скоростной «ручкой». Двухпедальные версии тоже есть: для 115-сильного дизеля предусмотрен 6-скоростной «автомат», для 115-сильного бензинового мотора — 7-скоростная роботизированная трансмиссия с двумя сцеплениями, для турбированного движка — вариатор.
Hyundai Creta в Индии можно купить с дизельными двигателями CRDi объёмом 1.4 и 1.6 литра, мощность которых составляет 90 и 128 л.с. соответственно. Также для кроссовера доступен 123-сильный бензиновый двигатель 1.6 литра. Версии с автоматической коробкой нет только у 1.4 CRDi.
Напомним, мировая премьера Kia Seltos состоялась 20 июня 2019 года в Дели. При этом в премьерном сообщении корейцы рассказали о силовых агрегатах для местного рынка, но не для индийского, где прошла презентация новинки.
Что касается моторной гаммы и вариантов трансмиссий для российского рынка, то в компании обещали рассказать подробности ближе к старту продаж, дата которого пока весьма абстрактна: Kia Seltos доберётся до РФ в 2020 году.
Фото ВАЗ 2115 1,3 | |||||
Модификация 2115 с двигателем 1,3 литра. | |||||
Характеристика ВАЗ 2115 1,3 | |||||
| Автомобиль ВАЗ | 2115 1,3 | ||||
| Кузов | |||||
| Тип кузова | седан | ||||
| Число мест | 5 | ||||
| Число дверей | 4 | ||||
| Габариты | |||||
| Длина, мм | 4330 | ||||
| Ширина, мм | 1620 | ||||
| Высота, мм | 1415 | ||||
| Колесная база, мм | 2460 | ||||
| Колея колес спереди, мм | 1400 | ||||
| Колея колес сзади, мм | 1370 | ||||
| Дорожный просвет, мм | 170 | ||||
| Шины | 165/70 R13 | ||||
| Снаряженная масса, кг | 970 | ||||
| Полная масса, кг | 1395 | ||||
| Полезная нагрузка, кг | 425 | ||||
| Объем багажника, л | 427 | ||||
| Объем топливного бака, л | 43 | ||||
| Двигатель | |||||
| Тип двигателя | L4 | ||||
| Объем двигателя, см³ | 1288 | ||||
Мощность, л. с./об.мин с./об.мин | 73/5600 | ||||
| Крутящий момент, Н·м/об.мин | 106/2800 | ||||
| Наддув | — | ||||
| Клапанов на цилиндр | 2 | ||||
| Расположение клапанов и распределительного вала | верхнеклапанный с верхним расположением распределительного вала | ||||
| Компоновка двигателя | спереди, поперечно | ||||
| Система питания | карбюратор | ||||
| Скорость | |||||
| Максимальная скорость, км/ч | 150 | ||||
| Разгон до 100 км/ч, с | 14,5 | ||||
| Топливо | |||||
| Марка топлива | бензин 92 | ||||
| Расход, л/100 км | 8,5 | ||||
| Привод | |||||
| Тип привода | передний | ||||
| КПП | |||||
| Механическая | 5 | ||||
| Автоматическая | — | ||||
| Подвеска | |||||
| Передняя | независимая типа Макферсон | ||||
| Задняя | продольный рычаг | ||||
| Тормоза | |||||
| Передние | дисковые | ||||
| Задние | барабанные | ||||
Размеры машины ваз 2115
Автомобили ВАЗ, 2113, 2114 и 2115 – модели, пользующиеся популярностью у отечественных водителей. Это связано с их невысокой стоимостью, ремонтопригодностью, относительной надежностью и неприхотливостью. А от объема багажника ВАЗ 2114 зависит практичность эксплуатации машины.
Это связано с их невысокой стоимостью, ремонтопригодностью, относительной надежностью и неприхотливостью. А от объема багажника ВАЗ 2114 зависит практичность эксплуатации машины.
ВАЗ 2113
Благодаря объему багажника ВАЗ 2113, у водителей есть возможность перевозить габаритные грузы. Его вместительность – 330 литров. Технические характеристики багажного отделения предоставляют возможность перевозить предметы, имеющие нестандартные размеры.
Багажник модели 2113 трансформируется: водитель может убрать полку, сложить задние сиденья. После этого его вместимость составляет 632 литра. Хэтчбек позволяет перевозить малогабаритные, длинномерные грузы.
К недостаткам относят отсутствие штатной подсветки (при тюнинге авто ее монтируют самостоятельно), высокий погрузочный бортик, который осложняет помещение грузов вовнутрь.
ВАЗ 2114
ВАЗ 2114 – надежный пятидверный автомобиль. Вместимость составляет 330 литров при сложенных сиденьях, а при разложенных – она равна 632 литрам. Погрузочная высота бортика имеет неудобный уровень (аналогично тому, который используется на тринадцатой модели), что существенно усложняет процесс погрузки.
Погрузочная высота бортика имеет неудобный уровень (аналогично тому, который используется на тринадцатой модели), что существенно усложняет процесс погрузки.
Габариты багажника выглядят следующим образом: длина – 830 миллиметров при сложенных сиденьях и практически 1660 миллиметров – при разложенных, ширина – 935 миллиметров, глубина – 830 миллиметров, высота – 780 миллиметров. Эти размеры приблизительные и актуальны для моделей 2113 и 2114.
Возможности трансформации позволяют загружать предметы, чьи длина, а также размеры отличаются от стандартных габаритов. При установке дополнительных багажных конструкций на крышу можно перевезти определенное количество крупногабаритных грузов.
Вместимость, выраженная в литрах, идентична объему, который указывается в кубических дециметрах.
ВАЗ 2115
Объем багажника ВАЗ 2115 составляет 427 литров при сложенных сиденьях, если их разложить, то вместимость увеличивается до 700 литров. Габариты багажного отделения увеличены, благодаря чему даже крупногабаритные грузы могут быть перевезены на пятнадцатой модели.
Габариты багажного отделения увеличены, благодаря чему даже крупногабаритные грузы могут быть перевезены на пятнадцатой модели.
К преимуществам относят простоту демонтажа, установку полки, а также удобную высоту для погрузки. Она находится практически на уровне автомобильного бампера.
Часто водители улучшают шумоизоляцию и штатную обшивку багажного отделения. Монтируют электрический привод открывания крышки, устанавливают подсветку, конструируют боксы для хранения небольших предметов (однако за счет их установки, багажник существенно теряет объем).
Особенности багажника
К особенностям багажных отделений различных модификаций моделей 2113, 2114, 2115 относят приемлемый уровень вместимости и возможность трансформации салона для увеличения полезного объема.
ВАЗ 2115 отличает удобная погрузочная высота, более увеличенный объем багажника Лада 2115. В остальном конструкция отделения для перевозки груза идентична другим моделям.
Авто оснащаются штатной полкой, она устанавливается вертикально и разделяет багажное отделение и салон. В случае необходимости (например, при перевозке крупногабаритных грузов, требующих складывания задних сидений), эту полку и вертикальную шторку можно демонтировать.
На автомобилях Лада 2113, 2114, 2115 под «полом» багажника предусмотрена ниша для хранения запасного колеса и инструментов, необходимых для быстрой починки небольших неисправностей, которые могут возникнуть в пути.
ВАЗ 2115 отличает удобная погрузочная высота, более увеличенный объем багажника Лада 2115. В остальном его конструкция идентична другим моделям.
Модели ВАЗ 2113, 2114, 2115, являющиеся модификацией «восьмерок», «девяток» и «девяносто девятых», соответственно, стали более эргономичными, удобными в использовании, практичными. Автомобили получили просторные багажные отделения, благодаря чему увеличилось количество перевозимого груза. Наличие ряда недостатков не отпугивает водителей: некоторые неудобства можно исправить самостоятельно.
Автомобили семейства ВАЗ 2113, 2114, 2115 популярны не только в России, но и в государствах СНГ. Это связано с большим распространением этих авто, их неприхотливостью, относительной надежностью, ремонтопригодностью и стоимостью. Машины обладают приемлемой вместимостью погрузочной ниши, что делает их удобными в использовании для решения повседневных бытовых задач или ведения небольшого бизнеса.
Кузов ВАЗ-2115 пришел на смену достаточно популярному у российских потребителей, но серьезно устаревшему корпусу 21099. Обще черты предшественницы без труда можно разглядеть у более новой модели.
В то же время 2115 получил немало и уникальных отличительных особенностей. К таковым следует отнести:
В этом материале будет рассказано, каковы размеры кузова ВАЗ-2115 и другие особенности его внешнего вида.
Изменения
В целом, невзирая на определенную схожесть, 2115 имеет значительно более современный дизайн. Во многом это произошло благодаря наличию бамперов и других внешних элементов, изготовленных из пластика. Все они позволяют защитить кузов от мелких повреждений и негативного действия коррозии.
Все они позволяют защитить кузов от мелких повреждений и негативного действия коррозии.
Наиболее заметные нововведения в этой модели следующие:
- добавочный тормозной сигнал, установленный непосредственно на спойлере;
- новые задние огни;
- высоконадежная система безопасности;
- порожек багажной двери, разрешивший снизить высоту погрузки;
- более обтекаемая геометрия кузова;
- молдинги на дверях;
- современное электрическое оборудование.
На момент завершения проектирования ВАЗ-2115 был современной и достаточно красивой машиной. При этом салон намного у нее оказался более комфортный, чем у старых моделей Волжского автозавода. Водитель получил возможность регулировать:
- высоту руля;
- положение сиденья и пр.
Между передними креслами конструкторы установили консоль. Более совершенной также оказалась и система обогрева, и ЭБУ. Причем багажник у рассматриваемой модели намного вместительней, чем у предшественниц.
Проектировщики немало элементов позаимствовали от 2114, при этом многие приборы и узлы перешли по наследству от «восьмерки». К примеру, неизменной перекочевала в новый седан ходовая часть.
Основные понятия
Геометрией кузова именуют точные размеры транспортного средства, установленные конструкторами завода-производителя. Основные параметры, имеющие первостепенное значение таковы:
- ширина – как передней, так и задней колей;
- протяженность колесной базы;
- расстояние между лонжеронами и их длина.
Как показывает практика, нарушение одного или сразу нескольких упомянутых выше параметров, приводит к заметному ухудшению в плане управляемости машины.
Второстепенные габариты следующие:
- проемы дверей;
- окон;
- объем отсека двигателя;
- вместительность багажника;
- размер салона машины.
Если происходит нарушение указанных нормативов, то это приводит только к ухудшению внешнего вида средства передвижения.
Колеей именуют расстояние, пролегающее между соосными колесами. Причем эта дистанция измеряется от точки до точки, которые расположены по центру протектора.
В свою очередь, колесной базой называют расстояние между осями автомобиля. Контрольные точки здесь – середина ступиц.
Составляющие части кузова и их размеры у ВАЗ-2115
Кузов в автомашине ВАЗ-2115 является несущим элементом всей конструкции. По типу данная модель считается 5-дверным седаном, приводимым в движение полуторалитровой силовой установкой.
Каркас кузова машины состоит из таких элементов:
- крыша;
- боковины;
- основание;
- неразборные части;
- навесные узлы;
- части интерьера.
В число навесных входят:
- капот;
- двери;
- крылья;
- бамперы;
- радиаторная декоративная решетка;
- крышка багажного отделения.
- длина автомобиля составляет в миллиметрах – 4330;
- от середины переднего колеса до передка – 860;
- от центра переднего до середины заднего колес – 2460;
- до задка от последней упомянутой точки – 1010;
- общая высота транспортного средства – 1420;
- дистанция между боковыми зеркалами – 1979;
- расстояние между колесами, расположенными спереди – 1400;
- габарит бампера, установленного сзади – 1650;
- задние колеса расположены друг от друга на 1370 миллиметров.

Рис. 1-1a. Габаритные размеры (справочные) автомобиля ВАЗ-2115
Рис. 1-1b. Габаритные размеры (справочные) автомобиля ВАЗ-2114
Ibanez представляет 15 новых привлекательных моделей RG к 2021 году
NAMM 2021 не происходит в физическом мире в этом году, но вы найдете все самые горячие январские выпуски оборудования в нашем путеводителе по самых крупных гитарных, усилителей и педальных релизов 2021 года на данный момент .
Серия RG компании Ibanez, производимая более трех десятилетий, является, возможно, краеугольным камнем ее продуктовой линейки. А к 2021 году японский гитарный гигант добавил в ассортимент широкий выбор новых моделей.
Девять новых электрогитар присоединяются к базовой серии RG, две модели присоединяются к серии RGA со скульптурным верхом, а линейка RGD, оптимизированная для дроп-мелодии, получает четыре новые модели, готовые к работе с рифами.
Ряд новых гитар может похвастаться хамбакерами Fishman Fluence, а линейка RGD отличается множеством потрясающих новых покрытий.
Эти модели присоединяются к трем новым дополнениям к серии AZ и новым гитарным гитарам Пола Гилберта, Пола Ваггонера и Мартина Миллера в модельном ряду компании 2021 года.
Без лишних слов, давайте углубимся в спецификации этих изящных новых топоров — вы можете найти больше информации на Ibanez.com.
RG Series
RG5320C
(Изображение предоставлено Ibanez)- Тип грифа: Super Wizard HP / 5 шт., Гриф из клена / венге
- Верх / спинка / корпус: Корпус из африканского красного дерева
- Гриф: Накладка грифа Macassar Ebony / офсетная инкрустация в виде точек из перламутра
- Лад: Лады Jumbo из нержавеющей стали / обработка кромки лада Prestige
- Количество ладов: 24
- Бридж: Lo-Pro Edge bridge
- Строка: 10. 8 мм
- Грифовый звукосниматель: Грифовый звукосниматель DiMarzio® Air Norton ™ (H) (пассивный / алнико)
- Бриджевый звукосниматель: DiMarzio® The Tone Zone® (H) бриджевый звукосниматель (пассивный / алнико)
- Factory настройка: 1E, 2B, 3G, 4D, 5A, 6E
- Струны: D’Addario® EXL120
- Калибр струны: .009 / .011 / .016 / .024 / .032 / .042
- Гайка: Стопорная гайка
- Цвет фурнитуры: Хром
- Органы управления: Регуляторы громкости и тона, трехпозиционный переключатель
 8 мм
8 ммRG5170B
(Изображение предоставлено: Ibanez)- Тип грифа : Super Wizard HP / 5шт клен / венге гриф
- Верх / спинка / корпус: Корпус из липы
- Гриф: Накладка на гриф из макассара черного дерева / вставка из перламутра
- Лад: Jumbo из нержавеющей стали Лады / Обработка кромки ладов Prestige
- Количество f rets: 24
- Bridge: Lo-Pro Edge bridge
- Струнное пространство: 10. 8 мм
- Нейловый звукосниматель: Нейловый звукосниматель Fishman® Fluence Modern Humbucker Alnico (H) (Active / Alnico)
- Средний звукосниматель: Fishman® Fluence Single Width Ceramic (S) средний звукосниматель (активный / керамический)
- Бриджевый звукосниматель: Бридж-звукосниматель Fishman® Fluence Modern Humbucker Ceramic (H) (активный / керамический)
- Заводская настройка: 1E, 2B, 3G, 4D, 5A, 6E
- Струны: D’Addario® EXL120
- Калибр струны: .009 / .011 / .016 / .024 / .032 / .042
- Гайка: Стопорная гайка
- Цвет фурнитуры: Золото (черные ручки)
- Элементы управления: Регуляторы громкости и тона, озвучка Fishman Fluence переключатель, пятипозиционный селекторный переключатель
 8 мм
8 ммRG5121
(Изображение предоставлено Ibanez)- Тип грифа: Super Wizard HP / 5 шт. Клен / венге, гриф
- Верх / спинка / корпус: Корпус из африканского красного дерева
- Гриф: Гриф из макассарского эбенового дерева / офсетная инкрустация в виде точек из перламутра
- Лад: Лады из нержавеющей стали Jumbo / обработка кромки лада Prestige
- Количество ладов: 24
- Бридж: Гибралтар Стандарт II мост
- Струнное пространство: 10.8 мм
- Нейловый звукосниматель: Нейловый звукосниматель Fishman® Fluence Modern Humbucker (H) (активный / керамический)
- Бриджевый звукосниматель: Бриджевый звукосниматель Fishman® Fluence Modern Humbucker (H) (активный / керамический)
- Заводская настройка : 1E, 2B, 3G, 4D, 5A, 6E
- Струны: D’Addario® EXL110
- Калибр струны: .010 / .013 / .017 / .026 / . 036 / .046
- Гайка: Graph Tech® BLACK Гайка TUSQ XL®
- Цвет фурнитуры: Cosmo black
- Элементы управления: Регуляторы громкости и тембра, переключатель голоса Fishman Fluence, трехпозиционный переключатель
 Клен / венге, гриф
Клен / венге, гриф 036 / .046
036 / .046RG752AHM
( Изображение предоставлено: Ibanez)- Тип грифа: Wizard-7/5 шт. Гриф из клена / венге
- Верх / спинка / корпус: Корпус из ясеня
- Гриф: Связанный клен Birdseye / вставка в черную точку
- Лад : Джамбо лады
- Бридж: Lo-P ro Edge 7 тремоло-бридж
- Струнное пространство: 10.8 мм
- Гриф: DiMarzio® PAF® 7 (H) гриф (пассивный / керамический)
- Бридж-звукосниматель: DiMarzio® PAF® 7 (H) бридж-звукосниматель (пассивный / керамический)
- Заводская настройка : 1E, 2B, 3G, 4D, 5A, 6E, 7B
- Калибр струны: . 009 / .011 / .016 / .024 / .032 / .042 / .054
- Цвет фурнитуры: Cosmo черный
- Органы управления: Регуляторы громкости и тона, пятипозиционный переключатель
 009 / .011 / .016 / .024 / .032 / .042 / .054
009 / .011 / .016 / .024 / .032 / .042 / .054RG652AHMFX
(Изображение предоставлено: Ibanez)- Тип грифа: Super Wizard HP / 5 шт. Гриф из клена / ореха
- Верх / задняя часть / корпус: Корпус из ясеня
- Гриф: Связанный клен Birdseye / инкрустация в виде черной точки
- Лад: Джамбо лад
- Бридж: Гибралтар Standard II бридж
- Струнное пространство: 10.8 мм
- Нейловый звукосниматель: Нейловый звукосниматель DiMarzio® Air Norton ™ (H) (Passive / Alnico)
- Бриджевый звукосниматель: DiMarzio® The Tone Zone® (H) бриджевый звукосниматель (Passive / Alnico)
- Factory настройка: 1E, 2B, 3G, 4D, 5A, 6E
- Калибр струны: . 010 / .013 / .017 / .026 / .036 / .046
- Гайка: Гайка Graph Tech®
- Цвет фурнитуры: Cosmo black
- Элементы управления: Ручки регулировки громкости и тона, пятипозиционный переключатель
 010 / .013 / .017 / .026 / .036 / .046
010 / .013 / .017 / .026 / .036 / .046RG6PKAG
(Изображение предоставлено Ibanez)- Тип грифа: Wizard / 11шт клен / орех / Гриф Bubinga
- Верх / спинка / корпус: Верх Koa / корпус из африканского красного дерева
- Гриф: Гриф из розового дерева / инкрустация в виде точек из перламутра
- Лад: Jescar EVO Gold лады / Обработка кромки лада Premium
- Количество ладов: 24
- Мост: Боковой мост
- Струнное пространство: 10.8 мм
- Гриф: EMG® SA (S) звукосниматель (Active / Alnico)
- Средний звукосниматель: EMG® SA (S) средний звукосниматель (Active / Alnico)
- Бридж-звукосниматель: EMG® 81 (H) бриджевый звукосниматель (активный / керамический)
- Заводская настройка: 1E, 2B, 3G, 4D, 5A, 6E
- Струны: D’Addario® EXL110
- Калибр струны: . 009 / .011 / .016 / .024 / .032 / .042
- Гайка: Стопорная гайка
- Цвет фурнитуры: Золото
 009 / .011 / .016 / .024 / .032 / .042
009 / .011 / .016 / .024 / .032 / .042Ibanez RG631ALF
(Изображение предоставлено: Ibanez)- Тип шеи: Nitro Wizard / 5pc Panga Panga / Гриф из орехового дерева
- Верх / спинка / корпус: Корпус Nyatoh
- Гриф: Макассар Эбеновый гриф / вставка с точечным смещением White Step
- Лад: Лады с обработкой Jumbo Sub Zero
- Количество ладов: 24
- Бридж: Гибралтар Стандарт II мост
- Струнное пространство: 10.8 мм
- Нейловый звукосниматель: Нейловый звукосниматель Fishman® Fluence Modern Humbucker Alnico (H) (Active / Alnico)
- Бридж-звукосниматель: Fishman® Fluence Modern Humbucker Ceramic (H) бридж-звукосниматель (активный / керамический)
- Заводская настройка: 1E, 2B, 3G, 4D, 5A, 6E
- Струны: D’Addario® EXL110
- Калибр струны: . 010 / .013 / .017 / .026 / .036 / .046
- Гайка: Пластиковая гайка
- Цвет фурнитуры: Черный
 010 / .013 / .017 / .026 / .036 / .046
010 / .013 / .017 / .026 / .036 / .046RG80F
(Изображение предоставлено: Ibanez)- Тип шеи: Wizard II-8/5 шт. Клен / гриф из ореха
- Верх / задняя часть / корпус: Корпус Meranti
- Гриф: Гриф из розового дерева / инкрустация в белые точки
- Лад: Джамбо лад
- Количество ладов: 24
- Бридж: F108 бридж
- Струнное пространство : 10.5 мм
- Нейловый звукосниматель: Нейловый звукосниматель Fishman® Fluence Modern Humbucker Alnico-8 (H) (Active / Alnico)
- Бридж-звукосниматель: Бридж-звукосниматель Fishman® Fluence Modern Humbucker Ceramic-8 (H) (активный / керамический )
- Заводская настройка: 1D #, 2A #, 3F #, 4C #, 5G #, 6D #, 7A #, 8F
- Струны: D’Addario® EXL120-7 + . 065
- Струны калибр: .009 / .011 / .016 / .024 / .032 / .042 / .054 / .065
- Гайка: Пластиковая гайка
- Цвет фурнитуры: Золото
 065
065RG320EXZ
( Изображение предоставлено: Ibanez)- Тип грифа: Wizard III Кленовый гриф
- Верх / спинка / корпус: Корпус Meranti
- Гриф: Связанный гриф Jatoba / вставка в белые точки
- Лад: Лады Jumbo
- Количество ладов: 24
- Бридж: Edge-Zero II с мостом ZPS3Fe
- String spa CE: 10.8 мм
- Нейловый звукосниматель: Нейловый звукосниматель Quantum (H) (пассивный / керамический)
- Бриджевый звукосниматель: бриджевый звукосниматель Quantum (H) (пассивный / керамический)
- Заводская настройка: 1E, 2B, 3G, 4D, 5A, 6E
- Струны: D’Addario® EXL120
- Калибр струны: . 009 / .011 / .016 / .024 / .032 / .042
- Гайка: Стопорная гайка
- Цвет фурнитуры: Черный
 009 / .011 / .016 / .024 / .032 / .042
009 / .011 / .016 / .024 / .032 / .042Серия RGA
RGA42HPT
(Изображение предоставлено Ibanez)- Тип грифа: Wizard III Жареный кленовый гриф
- Верх / спинка / корпус: Корпус Nyatoh
- Гриф: Гриф Jatoba / смещенная белая точка
- Лад: Джамбо лад
- Количество ладов: 24
- Бридж: Двойной замок тремоло-бридж
- Струнное пространство: 10.5 мм
- Гриф-звукосниматель: Гриф-звукосниматель DiMarzio® Fusion Edge (H) (пассивный / керамический)
- Бридж-звукосниматель: Бридж-звукосниматель DiMarzio® Fusion Edge (H) (пассивный / керамический)
- Заводская настройка: 1E, 2B, 3G, 4D, 5A, 6E
- Струны: D’Addario® EXL120
- Калибр струны: . 009 / .011 / .016 / .024 / .032 / .042
- Гайка : Стопорная гайка
- Цвет фурнитуры: Золото
 009 / .011 / .016 / .024 / .032 / .042
009 / .011 / .016 / .024 / .032 / .042RGA42HP
(Изображение предоставлено Ibanez)- Тип шеи: Wizard III Roasted Maple Neck
- Верх / спинка / корпус: Корпус Nyatoh
- Гриф: Гриф Jatoba / смещенная белая точка
- Лад: Джамбо лад
- Количество ладов: 24
- Бридж: F106 бридж
- Струнное пространство: 10.5 мм
- Гриф-звукосниматель: Гриф-звукосниматель DiMarzio® Fusion Edge (H) (пассивный / керамический)
- Бридж-звукосниматель: Бридж-звукосниматель DiMarzio® Fusion Edge (H) (пассивный / керамический)
- Заводская настройка: 1E, 2B, 3G, 4D, 5A, 6E
- Струны: D’Addario® EXL110
- Калибр струны: . 010 / .013 / .017 / .026 / .036 / .046
- Аппаратное обеспечение цвет: Gold
 010 / .013 / .017 / .026 / .036 / .046
010 / .013 / .017 / .026 / .036 / .046RGD Series
RGDR4427FX
(Изображение предоставлено: Ibanez)- Тип грифа: Wizard-7/5 шт. Клен / венге, гриф KTS ™ TITANIUM
- Верх / спинка / Корпус: Столешница Richlite / Корпус из ясеня
- Гриф: Гриф Bound Macassar Ebony / Смещенная инкрустация Luminlay dot
- Лад: Лады Jumbo / Обработка кромки ладов Prestige
- Количество ладов: 24
- Мост: Монорельсовый мост
- Расстояние между струнами: 10.8 мм
- Нейловый звукосниматель: Нейловый звукосниматель DiMarzio® Fusion Edge 7 (H) (пассивный / керамический)
- Бриджевый звукосниматель: Бриджевый звукосниматель DiMarzio® Fusion Edge 7 (H) (пассивный / керамический)
- Заводская настройка : 1D, 2A, 3F, 4C, 5G, 6D, 7A
- Струны: D’Addario® EXL110-7
- Калибр струны: . 010 / .013 / .017 / .026 / .036 / .046 / .059
- Гайка: Graph Tech® BLACK TUSQ XL® Гайка
- Цвет фурнитуры: Черный
 010 / .013 / .017 / .026 / .036 / .046 / .059
010 / .013 / .017 / .026 / .036 / .046 / .059RGD61ALA
(Изображение предоставлено Ibanez)- Тип шеи: Nitro Wizard 5 шт. Panga Panga / Гриф из орехового дерева
- Верх / спинка / корпус: Корпус из ясеня
- Гриф: Macassar Ebony Накладка с точечным смещением белого Step
- Лад: Лады с обработкой Jumbo Sub Zero
- Количество ладов: 24
- Мост: Монорельсовый мост
- Струнное пространство: 10.8 мм
- Нейловый звукосниматель: Нейловый звукосниматель Fishman® Fluence Modern Humbucker Alnico (H) (Active / Alnico)
- Бриджевый звукосниматель: Fishman® Fluence Modern Humbucker Ceramic (H) бриджевый звукосниматель (активный / керамический)
- Заводская настройка: 1D, 2A, 3F, 4C, 5G, 6D
- Струны: D’Addario® EXL110
- Калибр струны: . 010 / .013 / .017 / .026 / .036 / .046
- Гайка: Пластиковая гайка
- Цвет фурнитуры: Черный
 010 / .013 / .017 / .026 / .036 / .046
010 / .013 / .017 / .026 / .036 / .046RGD71ALPA
(Изображение предоставлено Ibanez)- Тип шеи: Nitro Wizard-7 / 5pc Panga Panga / Гриф из орехового дерева
- Верх / задняя часть / корпус: Тополь Тополь / корпус из многослойного ясеня и нято
- Гриф: Макассарское эбеновое дерево / вставка со смещенной точкой White Step
- Лад: Лады, обработанные Jumbo Sub Zero
- : 24
- Мост: Монорельс мост
- Струнное пространство: 10.8 мм
- Нейловый звукосниматель: Нейловый звукосниматель Bare Knuckle Aftermath-7 (H) (пассивный / Alnico)
- Бриджевый звукосниматель: Бридж-звукосниматель Bare Knuckle Aftermath-7 (H) (пассивный / керамический)
- Заводская настройка : 1D, 2A, 3F, 4C, 5G, 6D, 7A
- Струны: D’Addario® EXL110-7
- Калибр струны: . 010 / .013 / .017 / .026 / .036 / .046 / .059
- Гайка: Пластиковая гайка
- Цвет фурнитуры: Черный
 010 / .013 / .017 / .026 / .036 / .046 / .059
010 / .013 / .017 / .026 / .036 / .046 / .059RGD7521PB
(Изображение предоставлено Ibanez)- Тип грифа: Wizard II-7 Жареный кленовый гриф
- Верх / спинка / корпус: Верх из тополя / корпус Nyatoh
- Гриф: Гриф Ятоба / инкрустация в белые точки
- Лад: Джамбо лады
- Количество ладов: 24
- Мост: Гибралтар Стандартный мост II-7
- Струнное пространство: 10.8 мм
- Гриф: DiMarzio® PAF® 7 (H) гриф (пассивный / керамический)
- Бридж-звукосниматель: DiMarzio® PAF® 7 (H) бридж-звукосниматель (пассивный / керамический)
- Заводская настройка : 1D, 2A, 3F, 4C, 5G, 6D, 7A
- Струны: D’Addario® EXL110-7
- Калибр струны: . 010 / .013 / .017 / .026 / .036 / .046 / .059
- Гайка: Пластиковая гайка
- Цвет фурнитуры: Черный
 010 / .013 / .017 / .026 / .036 / .046 / .059
010 / .013 / .017 / .026 / .036 / .046 / .059Асимметричная локализация DLC1 определяет полярность нервного гребня птичьего ствола для направленного отслоения и миграции
Асимметричная активная RHOA указывает на миграционную поляризацию NC
Хотя было показано, что субклеточная локализация активности RHOA имеет решающее значение для определения полярности различных типов клеток 13 , В значительной степени неизвестно, участвует ли RHOA в установлении полярности ствола NC.Чтобы исследовать пространственно-временную динамику передачи сигналов RHOA в отслаивающихся и мигрирующих NCC, мы электропорировали каудальную нервную трубку куриных эмбрионов на стадиях Гамбургера и Гамильтона 14 (st) 11-12 с помощью одноцепочечного биосенсора на основе RHOA, который ранее использовался для обнаружить локальную активацию RHOA в фибробластах 15 . Через 24 часа после трансфекции (hpt) электропорированные эмбрионы на st 15-16 были собраны для FRET-анализа поперечного сечения грудной нервной трубки, где происходит отслоение и миграция NC.Визуализация FRET и измерение индекса FRET между задней и передней частью NCC выявили высокую активность RHOA в цитоплазме задней части клетки по сравнению с низкой или флуктуирующей активностью RHOA в выступах мембраны на переднем фронте расслаивающихся / рано- и поздно мигрирующих NCC. , которые отмечены спецификаторами NC, SOX9 и SOX10, соответственно (рис. 1a – d). Напротив, умеренный уровень активности RHOA обнаруживается и равномерно распределяется по нейроэпителиальным клеткам нервной трубки (дополнительный рис.1а, б). Эти результаты показывают, что NCC проявляют дифференциальную активность RHOA в субклеточной локализации, поскольку они подвергаются направленному отслаиванию и миграции. Чтобы изучить динамику активности RHOA в живых NCC, мы электропорировали эксплантаты нервной трубки с помощью биосенсора FRET и выполнили покадровую визуализацию NCC, подвергающихся направленному отслаиванию, на покрытые фибронектином чашки.
Через 24 часа после трансфекции (hpt) электропорированные эмбрионы на st 15-16 были собраны для FRET-анализа поперечного сечения грудной нервной трубки, где происходит отслоение и миграция NC.Визуализация FRET и измерение индекса FRET между задней и передней частью NCC выявили высокую активность RHOA в цитоплазме задней части клетки по сравнению с низкой или флуктуирующей активностью RHOA в выступах мембраны на переднем фронте расслаивающихся / рано- и поздно мигрирующих NCC. , которые отмечены спецификаторами NC, SOX9 и SOX10, соответственно (рис. 1a – d). Напротив, умеренный уровень активности RHOA обнаруживается и равномерно распределяется по нейроэпителиальным клеткам нервной трубки (дополнительный рис.1а, б). Эти результаты показывают, что NCC проявляют дифференциальную активность RHOA в субклеточной локализации, поскольку они подвергаются направленному отслаиванию и миграции. Чтобы изучить динамику активности RHOA в живых NCC, мы электропорировали эксплантаты нервной трубки с помощью биосенсора FRET и выполнили покадровую визуализацию NCC, подвергающихся направленному отслаиванию, на покрытые фибронектином чашки. В соответствии с наблюдениями in vivo, результаты визуализации показали, что активность RHOA была высоко обогащена в задней части клетки, а также динамически локализована в выступах мембраны на переднем крае поляризованных NCC, исходящих из эксплантов (рис.1e и дополнительный фильм 1). Чтобы определить постоянство активности RHOA, мы измерили индекс FRET между задней и передней частью поляризованных NCC с течением времени 4 . Мы обнаружили, что активность RHOA была постоянно высокой сзади, в то время как колебания уровней наблюдались спереди во время направленной миграции с течением времени (рис. 1f). Анализ общего сигнала FRET показал, что активность RHOA была выше сзади, чем спереди (рис. 1g). Кластеризация соотношения индексов FRET между задней и передней частью за 20 минут визуализации живых клеток подтвердила сохранение поляризованной активности RHOA во время миграции (рис.1ч). Чтобы дополнительно выяснить, сохраняется ли эта дифференциальная активность RHOA при изменении направления миграции NCC, эмигрирующие NCC из эксплантов нервной трубки подвергали воздействию гранул, покрытых фактором 1, полученным из стромальных клеток (SDF-1), который является хемоаттрактантом для NCC туловища 16 , чтобы имитировать среду in vivo.
В соответствии с наблюдениями in vivo, результаты визуализации показали, что активность RHOA была высоко обогащена в задней части клетки, а также динамически локализована в выступах мембраны на переднем крае поляризованных NCC, исходящих из эксплантов (рис.1e и дополнительный фильм 1). Чтобы определить постоянство активности RHOA, мы измерили индекс FRET между задней и передней частью поляризованных NCC с течением времени 4 . Мы обнаружили, что активность RHOA была постоянно высокой сзади, в то время как колебания уровней наблюдались спереди во время направленной миграции с течением времени (рис. 1f). Анализ общего сигнала FRET показал, что активность RHOA была выше сзади, чем спереди (рис. 1g). Кластеризация соотношения индексов FRET между задней и передней частью за 20 минут визуализации живых клеток подтвердила сохранение поляризованной активности RHOA во время миграции (рис.1ч). Чтобы дополнительно выяснить, сохраняется ли эта дифференциальная активность RHOA при изменении направления миграции NCC, эмигрирующие NCC из эксплантов нервной трубки подвергали воздействию гранул, покрытых фактором 1, полученным из стромальных клеток (SDF-1), который является хемоаттрактантом для NCC туловища 16 , чтобы имитировать среду in vivo. Мы наблюдали начальную поляризацию активности RHOA вдоль оси перед-назад, а последующая переназначение высокой RHOA в предполагаемую заднюю часть клетки была синхронизирована с образованием новых мембранных выступов на передней поверхности клетки, поскольку NCC подвергались изменению направления в ответ на SDF- 1 (рис.1i и j и дополнительный фильм 2). Количественная оценка индекса FRET между задней и передней частью во времени выявила поддержание высокой активности RHOA в задней части клетки, даже когда NCC изменяли направление своего движения (Fig. 1k). Таким образом, уже существующая асимметричная локализация активности RHOA определяет возможное направление поляризации клетки. Чтобы дополнительно коррелировать асимметричную активность RHOA с поляризацией клеток, мы исследовали динамику RHOA в популяции эмигрирующих NCC, которые претерпевают переключение вперед-назад в ответ на SDF-1.Покадровая визуализация FRET показала, что НК с поляризованной морфологией спереди-сзади постепенно приобретает повышенную активность RHOA спереди (A), которая в конечном итоге становится задней частью клетки после реполяризации клетки вместе с образованием нового выступа мембраны спереди, указывающего в сторону SDF-1 (B) (рис.
Мы наблюдали начальную поляризацию активности RHOA вдоль оси перед-назад, а последующая переназначение высокой RHOA в предполагаемую заднюю часть клетки была синхронизирована с образованием новых мембранных выступов на передней поверхности клетки, поскольку NCC подвергались изменению направления в ответ на SDF- 1 (рис.1i и j и дополнительный фильм 2). Количественная оценка индекса FRET между задней и передней частью во времени выявила поддержание высокой активности RHOA в задней части клетки, даже когда NCC изменяли направление своего движения (Fig. 1k). Таким образом, уже существующая асимметричная локализация активности RHOA определяет возможное направление поляризации клетки. Чтобы дополнительно коррелировать асимметричную активность RHOA с поляризацией клеток, мы исследовали динамику RHOA в популяции эмигрирующих NCC, которые претерпевают переключение вперед-назад в ответ на SDF-1.Покадровая визуализация FRET показала, что НК с поляризованной морфологией спереди-сзади постепенно приобретает повышенную активность RHOA спереди (A), которая в конечном итоге становится задней частью клетки после реполяризации клетки вместе с образованием нового выступа мембраны спереди, указывающего в сторону SDF-1 (B) (рис. 1l и m и дополнительный фильм 3). Количественная оценка активности FRET между A и B с течением времени показала, что перераспределение дифференциальной активности RHOA предшествовало началу постепенного переключения вперед и назад (рис.1н). В целом, исследования эксплантатов нервной трубки in vitro дополнительно консолидируют наблюдения in vivo о том, что существующая асимметрия активности RHOA иногда указывает на будущую обратную-переднюю полярность и направленную миграцию стволовых NCC.
1l и m и дополнительный фильм 3). Количественная оценка активности FRET между A и B с течением времени показала, что перераспределение дифференциальной активности RHOA предшествовало началу постепенного переключения вперед и назад (рис.1н). В целом, исследования эксплантатов нервной трубки in vitro дополнительно консолидируют наблюдения in vivo о том, что существующая асимметрия активности RHOA иногда указывает на будущую обратную-переднюю полярность и направленную миграцию стволовых NCC.
Асимметричная активность RHOA коррелирует с полярностью NC и направленной миграцией. Обработанные изображения сигнала FRET отслаивания a , ранней миграции b и поздней миграции NCC c . Увеличенная область и выбранные ячейки отмечены пунктирными квадратами.Белые стрелки указывают на сигнал FRET. Масштабные линейки 50 мкм. d Схематическое изображение асимметричной активности RHOA в поляризованном NCC. Количественная оценка соотношения индекса FRET между задней и передней частью отслаивающихся ( n = 107), ранними мигрирующими ( n = 123) и поздними мигрирующими NCC ( n = 196). Было проанализировано 25 эмбрионов. Среднее ± среднеквадратичное отклонение T-критерий Стьюдента, нс, не имеет значения. e Обработанные изображения сигнала FRET покадровой серии отслаивающихся NCC.Белые стрелки указывают на активность RHOA на фронте клетки. Масштабная линейка, 50 мкм. f Схематическое изображение асимметричной активности RHOA в поляризованном NCC, подвергающемся направленной миграции. Количественная оценка соотношения индекса FRET между задней и передней частью во времени. Было проанализировано n = 81 клетка из 26 эксплантов. г Количественная оценка общего индекса FRET в указанной области. Среднее ± s.e.m .; n = 81; Студенческий тест t ; *** р <0.0001. h Тепловые карты, представляющие индекс FRET как функцию времени сзади или спереди, и соотношение FRET между задней и передней частью 12 выбранных NCC из 12 эксплантов. i Обработанные изображения сигнала FRET покадровой серии мигрирующего NCC.
Было проанализировано 25 эмбрионов. Среднее ± среднеквадратичное отклонение T-критерий Стьюдента, нс, не имеет значения. e Обработанные изображения сигнала FRET покадровой серии отслаивающихся NCC.Белые стрелки указывают на активность RHOA на фронте клетки. Масштабная линейка, 50 мкм. f Схематическое изображение асимметричной активности RHOA в поляризованном NCC, подвергающемся направленной миграции. Количественная оценка соотношения индекса FRET между задней и передней частью во времени. Было проанализировано n = 81 клетка из 26 эксплантов. г Количественная оценка общего индекса FRET в указанной области. Среднее ± s.e.m .; n = 81; Студенческий тест t ; *** р <0.0001. h Тепловые карты, представляющие индекс FRET как функцию времени сзади или спереди, и соотношение FRET между задней и передней частью 12 выбранных NCC из 12 эксплантов. i Обработанные изображения сигнала FRET покадровой серии мигрирующего NCC. Белые стрелки указывают на изменение направления миграции в сторону SDF-1. Масштабная линейка, 50 мкм. j Схематическое изображение распределения активности RHOA в поляризованном NCC, претерпевающем изменение направления движения к шарикам, покрытым SDF-1. k Количественная оценка соотношения индекса FRET между задней и передней частью мигрирующих NCC. Было проанализировано n = 81 клетка из 21 экспланта. l Пример стационарного NCC, претерпевающего переключение вперед и назад в ответ на SDF-1. Было проанализировано n = 72 из 17 эксплантов. Белые стрелки указывают на начальное накопление высокой активности RHOA в передней части клетки. Белые стрелки указывают направление реполяризации клеток. Масштабная линейка, 50 мкм. м Схематическое изображение примера, показанного на рис.1л. n Количественная оценка отношения индекса FRET между A и B в примере, показанном на рис. 1l. Среднее ± s.e.m
Белые стрелки указывают на изменение направления миграции в сторону SDF-1. Масштабная линейка, 50 мкм. j Схематическое изображение распределения активности RHOA в поляризованном NCC, претерпевающем изменение направления движения к шарикам, покрытым SDF-1. k Количественная оценка соотношения индекса FRET между задней и передней частью мигрирующих NCC. Было проанализировано n = 81 клетка из 21 экспланта. l Пример стационарного NCC, претерпевающего переключение вперед и назад в ответ на SDF-1. Было проанализировано n = 72 из 17 эксплантов. Белые стрелки указывают на начальное накопление высокой активности RHOA в передней части клетки. Белые стрелки указывают направление реполяризации клеток. Масштабная линейка, 50 мкм. м Схематическое изображение примера, показанного на рис.1л. n Количественная оценка отношения индекса FRET между A и B в примере, показанном на рис. 1l. Среднее ± s.e.m
DLC1 асимметрично экспрессируется в мигрирующих NCC
В поисках генов, участвующих в регуляции поляризованной активности RHOA, мы идентифицировали белок, активирующий RhoGTPase (RhoGAP), удаленный при раке печени 1 (DLC1), который, как было показано, играет ключевую роль. в подавлении роста опухоли и метастазирования посредством подавления передачи сигналов RHO 17, 18 .У цыпленка есть три основные транскрипционные изоформы гена DLC1 (рис. 2а). Сравнительный анализ последовательностей показал, что их аминокислотные последовательности демонстрируют обширную общую гомологию с их аналогами как у человека, так и у мыши, с наибольшим сходством в четырех функциональных доменах, стерильном домене α-мотива (SAM), области нацеливания фокальной адгезии (FAT), RhoGAP и стероидогенный острый регуляторный домен (START) (дополнительный рис. 2a, b). Сначала мы исследовали экспрессию DLC1 в 8–14 куриных эмбрионах с использованием рибозонда против консервативного домена RhoGAP трех изоформ (рис.2а). DLC1 мРНК была первоначально обнаружена на st 8 и 9 в премигрирующих и расслаивающихся черепных NCC, которые коэкспрессируются со спецификатором NC, SOX9, и мигрирующим маркером NC, HNK-1 (Fig. 2d, e). DLC1 Экспрессия поддерживается в мигрирующих черепных NCCs в среднем мозге, областях заднего мозга и во фронтоназальном отростке от st 11-14 (Fig.
в подавлении роста опухоли и метастазирования посредством подавления передачи сигналов RHO 17, 18 .У цыпленка есть три основные транскрипционные изоформы гена DLC1 (рис. 2а). Сравнительный анализ последовательностей показал, что их аминокислотные последовательности демонстрируют обширную общую гомологию с их аналогами как у человека, так и у мыши, с наибольшим сходством в четырех функциональных доменах, стерильном домене α-мотива (SAM), области нацеливания фокальной адгезии (FAT), RhoGAP и стероидогенный острый регуляторный домен (START) (дополнительный рис. 2a, b). Сначала мы исследовали экспрессию DLC1 в 8–14 куриных эмбрионах с использованием рибозонда против консервативного домена RhoGAP трех изоформ (рис.2а). DLC1 мРНК была первоначально обнаружена на st 8 и 9 в премигрирующих и расслаивающихся черепных NCC, которые коэкспрессируются со спецификатором NC, SOX9, и мигрирующим маркером NC, HNK-1 (Fig. 2d, e). DLC1 Экспрессия поддерживается в мигрирующих черепных NCCs в среднем мозге, областях заднего мозга и во фронтоназальном отростке от st 11-14 (Fig. 2d). В заднем стволе от 11 до 13, DLC1 демонстрирует паттерн экспрессии, подобный SOX9 и другим спецификаторам NC, SOX10 и SNAIL2, в ранних мигрирующих NCC (рис.2г, е). Совпадая с градиентом развития NC от рострального к каудальному, DLC1 был сначала инициирован в премиграционной стадии и поддерживался в популяции мигрирующих черепных NC с последующей экспрессией в ранних мигрирующих NCC ствола. Чтобы дополнительно определить, какие изоформы экспрессируются в стволовых NCC, мы использовали энхансер SOX10 ( SOX10 -E1), чтобы управлять экспрессией репортера EGFP, специфически в вагусных / стволовых NCC 19 . Это позволило нам обогатить эту популяцию клеток с помощью сортировки клеток с активацией флуоресценции (FACS) для анализа количественной ПЦР с праймерами, специфичными для изоформ (рис.2б). Результаты показали, что изоформа 3 DLC1 была высоко экспрессирована в отсортированных клетках EGFP + по сравнению с низкими уровнями экспрессии изоформ DLC1 1 и 2 (рис.
2d). В заднем стволе от 11 до 13, DLC1 демонстрирует паттерн экспрессии, подобный SOX9 и другим спецификаторам NC, SOX10 и SNAIL2, в ранних мигрирующих NCC (рис.2г, е). Совпадая с градиентом развития NC от рострального к каудальному, DLC1 был сначала инициирован в премиграционной стадии и поддерживался в популяции мигрирующих черепных NC с последующей экспрессией в ранних мигрирующих NCC ствола. Чтобы дополнительно определить, какие изоформы экспрессируются в стволовых NCC, мы использовали энхансер SOX10 ( SOX10 -E1), чтобы управлять экспрессией репортера EGFP, специфически в вагусных / стволовых NCC 19 . Это позволило нам обогатить эту популяцию клеток с помощью сортировки клеток с активацией флуоресценции (FACS) для анализа количественной ПЦР с праймерами, специфичными для изоформ (рис.2б). Результаты показали, что изоформа 3 DLC1 была высоко экспрессирована в отсортированных клетках EGFP + по сравнению с низкими уровнями экспрессии изоформ DLC1 1 и 2 (рис. 2c), что указывает на то, что изоформа 3 преимущественно экспрессируется в магистральные НКЦ. Поэтому для простоты мы назвали изоформу 3 как DLC1 для остальных анализов.
2c), что указывает на то, что изоформа 3 преимущественно экспрессируется в магистральные НКЦ. Поэтому для простоты мы назвали изоформу 3 как DLC1 для остальных анализов.
Асимметричная локализация DLC1 отрицательно коррелирует с активностью RHOA. a Схематическое изображение трех изоформ DLC1 цыплят, содержащих четыре консервативных области: домен стерильного α-мотива (SAM), домен нацеливания на фокальную адгезию (FAT), домен, активирующий Rho GTPase (RhoGAP) и домен острой стероидогенной регуляции (START) . b Обогащенные проточной цитометрией NCC ствола (~ 1,62%), меченные SOX10 -управляемой энхансером (E1) экспрессией GFP. c уровней экспрессии мРНК изоформ DLC1 в отсортированных NCC. Среднее ± среднеквадратичное отклонение Тест Стьюдента t , *** p <0,0001 d Sense рибозонд для DLC1 служит отрицательным контролем. Гибридизация in situ DLC1 из куриных эмбрионов 8–14 стадий (st). Масштабная линейка 150 мкм. e Поперечный срез черепной области куриного эмбриона st 9, окрашенный DLC1 (черная линия) и иммунофлуоресценция для SOX9 и HNK-1 на последовательных срезах. f Поперечный срез в области туловища куриного эмбриона st 11, окрашенный DLC1 (черная линия) и иммунофлуоресценция для SOX9, SOX10 и SNAIL2 на последовательных срезах. Масштабные линейки, 50 мкм. Иммунофлуоресценция для DLC1 и фаллоидина при отслаивании g и ранних мигрирующих NCC h из эксплантатов нервной трубки и ядер окрашивают DAPI. Каналы DLC1 также показаны псевдоцветом. Белые пунктирные линии очерчивают границу эксплантата нервной пластинки. i , j Увеличение областей в рамке с повышенной экспрессией DLC1, показанной псевдоцветом. k Иммунофлуоресценция для FAK и DLC1 на мигрирующих NCC и ядрах окрашивают DAPI. -1 Анализ линейных сканирований, показывающий среднюю интенсивность флуоресценции DLC1 вдоль белой пунктирной линии за час .
Масштабная линейка 150 мкм. e Поперечный срез черепной области куриного эмбриона st 9, окрашенный DLC1 (черная линия) и иммунофлуоресценция для SOX9 и HNK-1 на последовательных срезах. f Поперечный срез в области туловища куриного эмбриона st 11, окрашенный DLC1 (черная линия) и иммунофлуоресценция для SOX9, SOX10 и SNAIL2 на последовательных срезах. Масштабные линейки, 50 мкм. Иммунофлуоресценция для DLC1 и фаллоидина при отслаивании g и ранних мигрирующих NCC h из эксплантатов нервной трубки и ядер окрашивают DAPI. Каналы DLC1 также показаны псевдоцветом. Белые пунктирные линии очерчивают границу эксплантата нервной пластинки. i , j Увеличение областей в рамке с повышенной экспрессией DLC1, показанной псевдоцветом. k Иммунофлуоресценция для FAK и DLC1 на мигрирующих NCC и ядрах окрашивают DAPI. -1 Анализ линейных сканирований, показывающий среднюю интенсивность флуоресценции DLC1 вдоль белой пунктирной линии за час . n = 47. m Репрезентативное изображение для измерения активности RHOA вдоль белой линии. Масштабные линейки, 20 мкм. n Количественная оценка активности RHOA ( n = 26) и o интенсивности флуоресценции DLC1 ( n = 47) от ведущего клеточного фронта к перинуклеарной области цитоплазмы
n = 47. m Репрезентативное изображение для измерения активности RHOA вдоль белой линии. Масштабные линейки, 20 мкм. n Количественная оценка активности RHOA ( n = 26) и o интенсивности флуоресценции DLC1 ( n = 47) от ведущего клеточного фронта к перинуклеарной области цитоплазмы
Поразительно, но иммунофлуоресцентный анализ эксплантатов нервной трубки показал, что DLC1 обнаруживает асимметричную локализацию в цитоплазме на фронте эмигрирующих и мигрирующих NCC (рис.2g – j), но не был локализован совместно с киназой фокальной адгезии (FAK), как показано в предыдущих исследованиях 20 (Fig. 2k). Сходным образом, когда меченый V5 DLC1 эктопически экспрессировался на низком уровне в нервной трубке цыплят, мы наблюдали преимущественную цитоплазматическую локализацию DLC1 впереди отслаивающихся и рано мигрирующих NCCs (Supplementary Fig. 3a). Чтобы количественно оценить асимметричную экспрессию DLC1, мы измерили интенсивность сигнала с помощью линейного сканирования вдоль тела клетки от различных выступов на переднем крае (рис. 2ч). Результаты показали, что экспрессия DLC1 была увеличена на фронте клетки в цитоплазме между выступами мембраны и ядром, тогда как экспрессия была низкой в ядре и сзади, что указывает на то, что DLC1 проявлял поляризованную экспрессию в начале отслоения NC (рис. 2l). . Затем мы сравнили паттерны совместной локализации между активностью RHOA и интенсивностью флуоресценции DLC1 от ведущего края клетки до перинуклеарной области цитоплазмы в разных группах клеток (рис. 2m) и подтвердили отрицательную корреляцию с высокой экспрессией DLC1 в области низкой RHOA. активность и наоборот (рис.2n, o), предполагая, что DLC1 может участвовать в установлении активности поляризованного RHOA в расслоении NCC.
2ч). Результаты показали, что экспрессия DLC1 была увеличена на фронте клетки в цитоплазме между выступами мембраны и ядром, тогда как экспрессия была низкой в ядре и сзади, что указывает на то, что DLC1 проявлял поляризованную экспрессию в начале отслоения NC (рис. 2l). . Затем мы сравнили паттерны совместной локализации между активностью RHOA и интенсивностью флуоресценции DLC1 от ведущего края клетки до перинуклеарной области цитоплазмы в разных группах клеток (рис. 2m) и подтвердили отрицательную корреляцию с высокой экспрессией DLC1 в области низкой RHOA. активность и наоборот (рис.2n, o), предполагая, что DLC1 может участвовать в установлении активности поляризованного RHOA в расслоении NCC.
DLC1 регулирует активность поляризованного RHOA и миграцию NC
Для дальнейшего изучения того, регулирует ли DLC1 пространственное ограничение активности RHOA для установления полярности NC и направленной миграции, мы сверхэкспрессировали биосенсор FRET вместе с DLC1, который обладает сильной активностью RhoGAP для ингибирования активной Форма экспрессии RHOA-GTP, определенная с помощью анализа активации RHOA pull-down, в каудальную геминуральную трубку куриных эмбрионов st 10–11 (рис. 3а, б). Через 24 часа анализ сигналов FRET на срезах показал, что активность RHOA была снижена или едва обнаруживалась в NCC, сверхэкспрессирующих DLC1, которые демонстрировали отсутствие оси полярности спереди назад (рис. 3c). Кроме того, сверхэкспрессия DLC1 не индуцирует эктопическую экспрессию SOX9, SOX10 и HNK-1 в нейроэпителии, указывая тем самым, что DLC1 недостаточно для запуска образования NC. Однако экспрессия этих маркеров в мигрирующих NCCs была снижена на трансфицированной стороне (Fig. 3e-h и u). Примечательно, что нейроэпителиальные клетки, сверхэкспрессирующие DLC1, отслаивались не только от базальной поверхности дорсальной нервной трубки, где был потерян ламинин, но также и в просвете (рис.3и, к). Соответственно, экспрессия N-Cadherin (N-Cad) теряется на апикальной поверхности дорсального нейроэпителия (Fig. 3k, l), указывая на то, что избыточная экспрессия DLC1 нарушает апикально-базальную полярность клеток дорсальной нервной трубки. Эти данные свидетельствуют о том, что высокий уровень экспрессии DLC1 снижает активность RHOA, что приводит к аберрантной полярности предмигрирующих NCC, которые отслаиваются в просвет нервной трубки вместо того, чтобы следовать своему нормальному маршруту миграции.
3а, б). Через 24 часа анализ сигналов FRET на срезах показал, что активность RHOA была снижена или едва обнаруживалась в NCC, сверхэкспрессирующих DLC1, которые демонстрировали отсутствие оси полярности спереди назад (рис. 3c). Кроме того, сверхэкспрессия DLC1 не индуцирует эктопическую экспрессию SOX9, SOX10 и HNK-1 в нейроэпителии, указывая тем самым, что DLC1 недостаточно для запуска образования NC. Однако экспрессия этих маркеров в мигрирующих NCCs была снижена на трансфицированной стороне (Fig. 3e-h и u). Примечательно, что нейроэпителиальные клетки, сверхэкспрессирующие DLC1, отслаивались не только от базальной поверхности дорсальной нервной трубки, где был потерян ламинин, но также и в просвете (рис.3и, к). Соответственно, экспрессия N-Cadherin (N-Cad) теряется на апикальной поверхности дорсального нейроэпителия (Fig. 3k, l), указывая на то, что избыточная экспрессия DLC1 нарушает апикально-базальную полярность клеток дорсальной нервной трубки. Эти данные свидетельствуют о том, что высокий уровень экспрессии DLC1 снижает активность RHOA, что приводит к аберрантной полярности предмигрирующих NCC, которые отслаиваются в просвет нервной трубки вместо того, чтобы следовать своему нормальному маршруту миграции.
DLC1 пространственно ограничивает активность RHOA и регулирует миграционное поведение NC. a Схематическая диаграмма полноразмерного DLC1 и доминантно-отрицательного DLC1 (DN-DLC1), содержащих усеченный на С-конце фрагмент DLC1 без доменов RhoGAP и START. b Иммуноблоттинг для RHOA-GTP на лизатах белков, экстрагированных из нервных трубок, подвергнутых электропорации с указанными конструкциями через 24 ч. Общая RHOA используется в качестве контроля загрузки. Столбцы представляют результаты денситометрического анализа (среднее ± стандартное отклонение, n = 3 независимых эксперимента). Студенческий t -тест, * p <0.05; нс, не имеет значения. c Обработанные изображения сигналов FRET поперечных сечений эмбрионов, подвергнутых электропорации с помощью датчика FRET плюс DLC1 или DN-DLC1. NCC маркируются иммунофлуоресценцией SOX9, а ядра окрашиваются DAPI. Увеличенная область и выбранные ячейки отмечены пунктирными квадратами. Белые стрелки указывают на сигнал FRET. Масштабные линейки 20 мкм. d Количественная оценка общего индекса FRET в контроле ( n = 425/25 эмбрионов), NCC, экспрессирующих DN-DLC1 ( n = 148/21 эмбрионов) и DLC1 ( n = 131/16 эмбрионов).Студенческий t -тест, ** p <0,001; *** р <0,0001. e — l Иммунофлуоресценция для SOX9, SOX10, HNK-1, ламинина и N-Cad на поперечных срезах эмбрионов, подвергнутых электропорации (EP) с DLC1 на 24 hpt ( n = 12). На вставках показаны объединенные изображения GFP и эндогенной экспрессии SOX9 или SOX10. Белый треугольник указывает на экспрессию эндогенного HNK-1 в мигрирующих NCC, тогда как белые треугольники указывают на снижение экспрессии HNK-1, ламинина и N-Cad на трансфицированной стороне нервной трубки.Масштабные линейки, 50 мкм. m , n Иммунофлуоресценция для HNK-1 в эмбрионе, подвергнутом электропорации с DN-DLC1 через 24 часа ( n = 17). Масштабная линейка 150 мкм.
Белые стрелки указывают на сигнал FRET. Масштабные линейки 20 мкм. d Количественная оценка общего индекса FRET в контроле ( n = 425/25 эмбрионов), NCC, экспрессирующих DN-DLC1 ( n = 148/21 эмбрионов) и DLC1 ( n = 131/16 эмбрионов).Студенческий t -тест, ** p <0,001; *** р <0,0001. e — l Иммунофлуоресценция для SOX9, SOX10, HNK-1, ламинина и N-Cad на поперечных срезах эмбрионов, подвергнутых электропорации (EP) с DLC1 на 24 hpt ( n = 12). На вставках показаны объединенные изображения GFP и эндогенной экспрессии SOX9 или SOX10. Белый треугольник указывает на экспрессию эндогенного HNK-1 в мигрирующих NCC, тогда как белые треугольники указывают на снижение экспрессии HNK-1, ламинина и N-Cad на трансфицированной стороне нервной трубки.Масштабные линейки, 50 мкм. m , n Иммунофлуоресценция для HNK-1 в эмбрионе, подвергнутом электропорации с DN-DLC1 через 24 часа ( n = 17). Масштабная линейка 150 мкм. o , p Поперечное сечение на уровне белой линии в м и n. Белые треугольники указывают на экспрессию эндогенного HNK-1 в мигрирующих NCC, тогда как белые треугольники указывают на снижение экспрессии HNK-1 на трансфицированной стороне. q — t Иммунофлуоресценция для SOX9, SOX10, N-Cad и ламинина на поперечных срезах эмбрионов, электропорированных с помощью DN-DLC1 на 24 hpt ( n = 17).На вставках показаны объединенные изображения зеленого и красного каналов. Масштабная линейка, 50 мкм. и Количественная оценка количества эмигрирующих клеток GFP + , экспрессирующих HNK-1, на трансфицированной стороне нервной трубки, подвергнутой электропорации с указанными конструкциями. Среднее количество клеток, подсчитанных по крайней мере из 30 срезов 10 эмбрионов за обработку. Среднее ± среднеквадратичное отклонение Студенческий т -тест ** р <0,001
o , p Поперечное сечение на уровне белой линии в м и n. Белые треугольники указывают на экспрессию эндогенного HNK-1 в мигрирующих NCC, тогда как белые треугольники указывают на снижение экспрессии HNK-1 на трансфицированной стороне. q — t Иммунофлуоресценция для SOX9, SOX10, N-Cad и ламинина на поперечных срезах эмбрионов, электропорированных с помощью DN-DLC1 на 24 hpt ( n = 17).На вставках показаны объединенные изображения зеленого и красного каналов. Масштабная линейка, 50 мкм. и Количественная оценка количества эмигрирующих клеток GFP + , экспрессирующих HNK-1, на трансфицированной стороне нервной трубки, подвергнутой электропорации с указанными конструкциями. Среднее количество клеток, подсчитанных по крайней мере из 30 срезов 10 эмбрионов за обработку. Среднее ± среднеквадратичное отклонение Студенческий т -тест ** р <0,001
Чтобы проверить, влияет ли ингибирование функции DLC1 на активность RHOA, судьбу, полярность и миграционное поведение NCC, мы создали доминантно-отрицательную версию DLC1 (DN-DLC1), состоящую из аминоконцевой области DLC1 без доменов RhoGAP и START ( Инжир. 3а). Ранее было показано, что эта конструкция нарушает ассоциацию DLC1 с его партнерским фактором, что приводит к ингибированию подвижности клеток in vitro 21 . В отличие от DLC1, сверхэкспрессия DN-DLC1 приводит к увеличению уровня RHOA-GTP (рис. 3b). Чтобы дополнительно продемонстрировать прямое ингибирующее действие DN-DLC1 на DLC1, мы совместно электропорировали DN-DLC1 с DLC1 и обнаружили, что уровень RHOA-GTP был немного снижен по сравнению с контролем и DLC1 (рис. 3b), что указывает на то, что DN- DLC1 действительно действует как доминантно-негативный, ингибируя эндогенную активность RhoGAP DLC1.Соответственно, сверхэкспрессия DN-DLC1 in vivo приводит к заметному повышению активности RHOA, которая равномерно распределяется по цитоплазме расслаивающихся NCCs без четкой субклеточной локализации, и клетки имеют округлую форму (Fig. 3c). Количественный анализ общей активности FRET in vivo показал, что по сравнению с векторным контролем общие уровни активации RHOA были значительно увеличены (тест Стьюдента t , p <0,0001) и уменьшились (тест Стьюдента t , р <0.
3а). Ранее было показано, что эта конструкция нарушает ассоциацию DLC1 с его партнерским фактором, что приводит к ингибированию подвижности клеток in vitro 21 . В отличие от DLC1, сверхэкспрессия DN-DLC1 приводит к увеличению уровня RHOA-GTP (рис. 3b). Чтобы дополнительно продемонстрировать прямое ингибирующее действие DN-DLC1 на DLC1, мы совместно электропорировали DN-DLC1 с DLC1 и обнаружили, что уровень RHOA-GTP был немного снижен по сравнению с контролем и DLC1 (рис. 3b), что указывает на то, что DN- DLC1 действительно действует как доминантно-негативный, ингибируя эндогенную активность RhoGAP DLC1.Соответственно, сверхэкспрессия DN-DLC1 in vivo приводит к заметному повышению активности RHOA, которая равномерно распределяется по цитоплазме расслаивающихся NCCs без четкой субклеточной локализации, и клетки имеют округлую форму (Fig. 3c). Количественный анализ общей активности FRET in vivo показал, что по сравнению с векторным контролем общие уровни активации RHOA были значительно увеличены (тест Стьюдента t , p <0,0001) и уменьшились (тест Стьюдента t , р <0. 001) в NCC, сверхэкспрессирующих DN-DLC1 и DLC1, соответственно (Fig. 3d). Анализ маркеров NC показал, что избыточная экспрессия DN-DLC1 приводит к заметно меньшему количеству мигрирующих NCCs, экспрессирующих HNK-1, по сравнению с контралатеральной стороной (Fig. 3m – p и u). Соответственно, количество ранних мигрирующих NCC, экспрессирующих SOX9 и SOX10, было снижено на трансфицированной стороне, но их экспрессия в премиграторном домене осталась неизменной, что указывает на то, что на идентичность NC не повлияла избыточная экспрессия DN-DLC1 (рис.3q, r). В отличие от DLC1, большинство NCCs, экспрессирующих DN-DLC1, остается в нейроэпителии без нарушения апикально-базальной полярности (Fig. 3s, t). Чтобы дополнительно продемонстрировать, что DLC1 необходим для полярности NC и расслоения, мы электропорировали меченную флуоресцеином морфолино, нацеленную на изоформу 3 DLC1 (DLC1-MO), в каудальную геминеральную трубку эмбрионов st 10-11. Через 24 часа белок DLC1 уменьшился при использовании более высокой дозировки DLC1-MO, тогда как его экспрессия оставалась неизменной у эмбрионов, обработанных только контрольным-MO (Ctrl-MO) и вектором (дополнительный рис.
001) в NCC, сверхэкспрессирующих DN-DLC1 и DLC1, соответственно (Fig. 3d). Анализ маркеров NC показал, что избыточная экспрессия DN-DLC1 приводит к заметно меньшему количеству мигрирующих NCCs, экспрессирующих HNK-1, по сравнению с контралатеральной стороной (Fig. 3m – p и u). Соответственно, количество ранних мигрирующих NCC, экспрессирующих SOX9 и SOX10, было снижено на трансфицированной стороне, но их экспрессия в премиграторном домене осталась неизменной, что указывает на то, что на идентичность NC не повлияла избыточная экспрессия DN-DLC1 (рис.3q, r). В отличие от DLC1, большинство NCCs, экспрессирующих DN-DLC1, остается в нейроэпителии без нарушения апикально-базальной полярности (Fig. 3s, t). Чтобы дополнительно продемонстрировать, что DLC1 необходим для полярности NC и расслоения, мы электропорировали меченную флуоресцеином морфолино, нацеленную на изоформу 3 DLC1 (DLC1-MO), в каудальную геминеральную трубку эмбрионов st 10-11. Через 24 часа белок DLC1 уменьшился при использовании более высокой дозировки DLC1-MO, тогда как его экспрессия оставалась неизменной у эмбрионов, обработанных только контрольным-MO (Ctrl-MO) и вектором (дополнительный рис. 4а). Подобное снижение экспрессии DLC1 наблюдалось в обработанных DLC1-MO культивируемых NCC, которые имели округлую форму без отчетливой передне-задней полярности, как клетки, экспрессирующие DN-DLC1 (дополнительные фиг. 4b и 3c). В соответствии с этим, клетки, экспрессирующие DLC1-MO, приводили к снижению экспрессии FOXD3 + мигрирующих NCC по сравнению с нетрансфицированной стороной и эмбрионами, обработанными Ctrl-MO (дополнительный рис. 4c). Эти результаты демонстрируют функциональные требования к DLC1 в установлении полярности NC и расслоения.В целом эти исследования in vivo показывают, что соответствующий уровень активности DLC1 необходим для пространственного ограничения активности RHOA, установления полярности NC и направленного отслоения NCC.
4а). Подобное снижение экспрессии DLC1 наблюдалось в обработанных DLC1-MO культивируемых NCC, которые имели округлую форму без отчетливой передне-задней полярности, как клетки, экспрессирующие DN-DLC1 (дополнительные фиг. 4b и 3c). В соответствии с этим, клетки, экспрессирующие DLC1-MO, приводили к снижению экспрессии FOXD3 + мигрирующих NCC по сравнению с нетрансфицированной стороной и эмбрионами, обработанными Ctrl-MO (дополнительный рис. 4c). Эти результаты демонстрируют функциональные требования к DLC1 в установлении полярности NC и расслоения.В целом эти исследования in vivo показывают, что соответствующий уровень активности DLC1 необходим для пространственного ограничения активности RHOA, установления полярности NC и направленного отслоения NCC.
DLC1 пространственно ограничивает RHOA для направленной миграции NC
Для дальнейшей оценки того, как изменение активности DLC1 более подробно влияет на динамику миграционного поведения NC, мы выполнили покадровую визуализацию in vitro отслаивающихся NCC от эксплантатов нервной трубки, электропорированных вектором. контроль, DLC1 и DN-DLC1 и исследовали эффекты клеточной полярности на основе степени выступов мембраны 22, 23 .Поскольку образование мембранных выступов управляется динамикой актина, NCCs также трансфицировали Lifeact-mCherry для маркировки актинового цитоскелета для количественной оценки углов между выступом мембраны и направлением миграции (Fig. 4a). Как и ожидалось, клетки, экспрессирующие GFP, мигрировали из эксплантатов и демонстрировали характерный образец мембранных выступов на клеточном фронте в направлении миграции (Fig. 4a и Supplementary Movie 4). Напротив, NCC, сверхэкспрессирующие DLC1, теряли переднюю-заднюю поляризацию со случайно ориентированными и беспорядочными пальцеобразными выступами и демонстрировали потерю направленности (рис.4а и дополнительный фильм 5). Мембранные выступы в форме пузырей наблюдались в клетках, экспрессирующих DN-DLC1, которые проявляли ограниченную подвижность (рис. 4а и дополнительный ролик 6). Затем мы измерили набор параметров миграции, чтобы определить, как изменение полярности NC путем манипулирования активностью DLC1 влияет на миграционное поведение.
контроль, DLC1 и DN-DLC1 и исследовали эффекты клеточной полярности на основе степени выступов мембраны 22, 23 .Поскольку образование мембранных выступов управляется динамикой актина, NCCs также трансфицировали Lifeact-mCherry для маркировки актинового цитоскелета для количественной оценки углов между выступом мембраны и направлением миграции (Fig. 4a). Как и ожидалось, клетки, экспрессирующие GFP, мигрировали из эксплантатов и демонстрировали характерный образец мембранных выступов на клеточном фронте в направлении миграции (Fig. 4a и Supplementary Movie 4). Напротив, NCC, сверхэкспрессирующие DLC1, теряли переднюю-заднюю поляризацию со случайно ориентированными и беспорядочными пальцеобразными выступами и демонстрировали потерю направленности (рис.4а и дополнительный фильм 5). Мембранные выступы в форме пузырей наблюдались в клетках, экспрессирующих DN-DLC1, которые проявляли ограниченную подвижность (рис. 4а и дополнительный ролик 6). Затем мы измерили набор параметров миграции, чтобы определить, как изменение полярности NC путем манипулирования активностью DLC1 влияет на миграционное поведение. Расстояние перемещения, устойчивость и чистая скорость миграции клеток были уменьшены как в NCC, сверхэкспрессирующих DLC1, так и в DN-DLC1, по сравнению с векторным контролем и клетками, не экспрессирующими GFP (рис.4b – e), что указывает на отсутствие направленности. Общая скорость миграции NCC, экспрессирующих DN-DLC1, была заметно ниже, чем скорость миграции клеток со сверхэкспрессией DLC1, которые мигрировали со сравнимой скоростью с контрольными и не экспрессирующими GFP клетками (фиг. 4f). Эти результаты показывают, что сверхэкспрессия и доминантно-отрицательное ингибирование активности DLC1 нарушают поляризацию NCC и направленную миграцию. Затем мы оценили их влияние на активность RHOA в культивируемых NCC, трансфицированных FRET-биосенсором в присутствии SDF-1, чтобы определить, можно ли восстановить поляризованную морфологию и миграционное поведение NCC с помощью управляющего сигнала (рис.4г). В соответствии с наблюдениями in vivo, покадровая визуализация и индекс FRET между задней и передней частью показали, что сверхэкспрессия DLC1 и DN-DLC1 приводила к заметному снижению и повышению активности RHOA во всей клетке соответственно по сравнению с векторным контролем, который демонстрировал воспроизводимое поляризованное распределение активности RHOA со стойким высоким сигналом FRET сзади и мигрировало в сторону SDF-1 (рис.
Расстояние перемещения, устойчивость и чистая скорость миграции клеток были уменьшены как в NCC, сверхэкспрессирующих DLC1, так и в DN-DLC1, по сравнению с векторным контролем и клетками, не экспрессирующими GFP (рис.4b – e), что указывает на отсутствие направленности. Общая скорость миграции NCC, экспрессирующих DN-DLC1, была заметно ниже, чем скорость миграции клеток со сверхэкспрессией DLC1, которые мигрировали со сравнимой скоростью с контрольными и не экспрессирующими GFP клетками (фиг. 4f). Эти результаты показывают, что сверхэкспрессия и доминантно-отрицательное ингибирование активности DLC1 нарушают поляризацию NCC и направленную миграцию. Затем мы оценили их влияние на активность RHOA в культивируемых NCC, трансфицированных FRET-биосенсором в присутствии SDF-1, чтобы определить, можно ли восстановить поляризованную морфологию и миграционное поведение NCC с помощью управляющего сигнала (рис.4г). В соответствии с наблюдениями in vivo, покадровая визуализация и индекс FRET между задней и передней частью показали, что сверхэкспрессия DLC1 и DN-DLC1 приводила к заметному снижению и повышению активности RHOA во всей клетке соответственно по сравнению с векторным контролем, который демонстрировал воспроизводимое поляризованное распределение активности RHOA со стойким высоким сигналом FRET сзади и мигрировало в сторону SDF-1 (рис. 4h, i и дополнительные видеоролики 7–9). Кроме того, мы наблюдали аналогичное отсутствие полярности клеток и направленного движения в обоих вариантах лечения, предполагая, что добавление SDF-1 в культуру эксплантата нервной трубки не способно восстановить морфологические и миграционные дефекты, вызванные аберрантной передачей сигналов RHOA (рис.4ч). В целом, эти исследования in vitro дополнительно консолидируют результаты in vivo о том, что для расслоения NCC требуется точная пространственная активность DLC1 для ограничения высокой активности RHOA в спине, что важно для приобретения морфологии поляризованных NCC и подвижности к источнику хемоаттрактанта.
4h, i и дополнительные видеоролики 7–9). Кроме того, мы наблюдали аналогичное отсутствие полярности клеток и направленного движения в обоих вариантах лечения, предполагая, что добавление SDF-1 в культуру эксплантата нервной трубки не способно восстановить морфологические и миграционные дефекты, вызванные аберрантной передачей сигналов RHOA (рис.4ч). В целом, эти исследования in vitro дополнительно консолидируют результаты in vivo о том, что для расслоения NCC требуется точная пространственная активность DLC1 для ограничения высокой активности RHOA в спине, что важно для приобретения морфологии поляризованных NCC и подвижности к источнику хемоаттрактанта.
Соответствующий уровень активности DLC1 необходим для направленной миграции NCC. a Покадровая съемка, показывающая динамику выпячивания эмигрирующих NCC, выражающих контроль GFP, DLC1 и DN-DLC1.Белые стрелки указывают направление выступов. Полярные гистограммы, показывающие распределение угла между выступами актина и направление последующей миграции для всех временных рядов. n = 22/10 эксплантов для контроля GFP, n = 20/12 эксплантов для DLC1, n = 25/12 эксплантов для DN-DLC1. Масштабная линейка 20 мкм. b Показаны следы миграции 17 репрезентативных NCC, выражающих контроль GFP, DN-DLC1 и DLC1. Количественная оценка общего расстояния миграции c , персистенции d , чистой скорости e и общей скорости f NCC, подвергнутых электропорации с указанными конструкциями.GFP и не-GFP клетки служат в качестве контроля. Среднее ± s.e.m. Студенческий t -тест, *** p <0,0001; ** p <0,001; нс, не имеет значения. g Схематическая диаграмма, показывающая заднюю (сторона B) и переднюю сторону (сторона A) клетки нервного гребня, мигрирующей к шарикам, покрытым SDF-1. Масштабная линейка 50 мкм. ч Обработанные изображения сигнала FRET покадровой серии эмигрирующих NCC из эксплантатов нервной трубки, обработанных векторным контролем ( n = 111/16 эксплантатов), DLC1 ( n = 93/17 эксплантов) и DN-DLC1 ( n = 88/13 эксплантов).
n = 22/10 эксплантов для контроля GFP, n = 20/12 эксплантов для DLC1, n = 25/12 эксплантов для DN-DLC1. Масштабная линейка 20 мкм. b Показаны следы миграции 17 репрезентативных NCC, выражающих контроль GFP, DN-DLC1 и DLC1. Количественная оценка общего расстояния миграции c , персистенции d , чистой скорости e и общей скорости f NCC, подвергнутых электропорации с указанными конструкциями.GFP и не-GFP клетки служат в качестве контроля. Среднее ± s.e.m. Студенческий t -тест, *** p <0,0001; ** p <0,001; нс, не имеет значения. g Схематическая диаграмма, показывающая заднюю (сторона B) и переднюю сторону (сторона A) клетки нервного гребня, мигрирующей к шарикам, покрытым SDF-1. Масштабная линейка 50 мкм. ч Обработанные изображения сигнала FRET покадровой серии эмигрирующих NCC из эксплантатов нервной трубки, обработанных векторным контролем ( n = 111/16 эксплантатов), DLC1 ( n = 93/17 эксплантов) и DN-DLC1 ( n = 88/13 эксплантов). Масштабные линейки 50 мкм. i Количественное определение соотношения индекса FRET между задней (B) и передней (A) частью кластера для указанных условий. Среднее ± s.e.m. Студенческий т -тест, *** р <0,0001
Масштабные линейки 50 мкм. i Количественное определение соотношения индекса FRET между задней (B) и передней (A) частью кластера для указанных условий. Среднее ± s.e.m. Студенческий т -тест, *** р <0,0001
NEDD9 необходим для асимметричной локализации DLC1
Предыдущие исследования показали, что DLC1 взаимодействует с другими факторами, чтобы модулировать его функцию RhoGAP 17 . Чтобы идентифицировать белковые факторы, связанные с DLC1, мы выполнили иммунопреципитацию антителом DLC1, специфичным для изоформы 3, с использованием лизатов белка нервной трубки цыплят.Протеомный анализ выявил различные функциональные категории белков, обогащенных DLC1 (рис. 5a, дополнительные рис. 5a, b и дополнительные данные 1). Анализ базы знаний Ingenuity Pathway показал, что большинство этих белков участвуют в передаче сигналов EIF2, ремоделировании эпителиальных адгезивных соединений и регуляции основанной на актине подвижности с помощью RHO (Fig.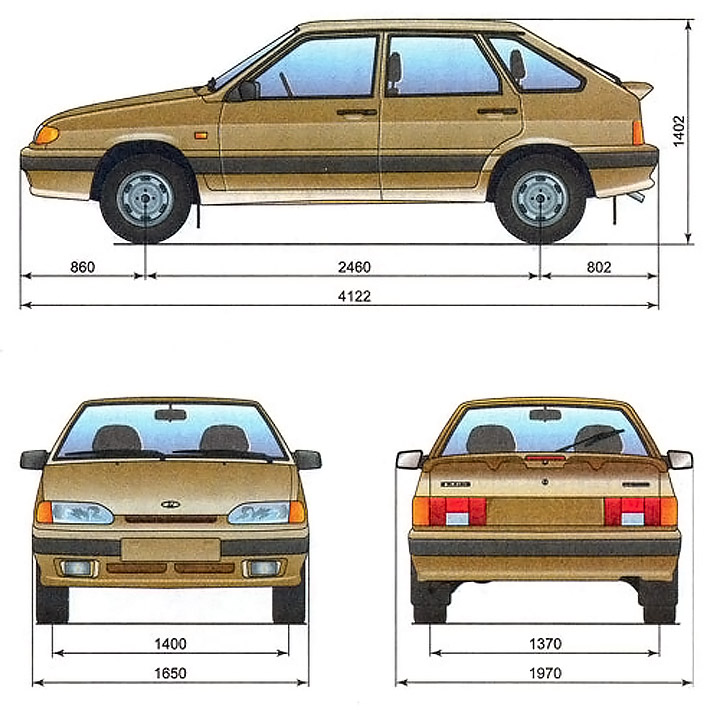 5b and Supplementary Data 2). Соответственно, сеть интерактомов показала, что белки, относящиеся к клеточной подвижности и регуляции цитоскелета, были в высокой степени связаны с DLC1 (рис.5в). NEDD9, член семейства каркасных белков Cas, был выбран для дальнейшего исследования, поскольку было показано, что он участвует в подвижности NC 24 , хотя механизм действия неизвестен. Коиммунопреципитация подтвердила ассоциацию эндогенных белков DLC1 и NEDD9 в эмбриональных нервных трубках (рис. 5d), что повышает вероятность того, что DN-DLC1 может мешать взаимодействию DLC1-NEDD9, исходя из способа его действия из предыдущего исследования 21 . В самом деле, эктопическая экспрессия DN-DLC1 также способна коиммунопреципитировать с NEDD9.Дальнейшее картирование доменов показало, что усеченная на С-конце версия DN-DLC1, состоящая из доменов SAM и FAT [DN-DLC1 (320)] 25 , где большинство партнеров по взаимодействию связывания DLC1 могут связываться с NEDD9, тогда как домен SAM сам по себе не показать взаимодействие с NEDD9 (рис.
5b and Supplementary Data 2). Соответственно, сеть интерактомов показала, что белки, относящиеся к клеточной подвижности и регуляции цитоскелета, были в высокой степени связаны с DLC1 (рис.5в). NEDD9, член семейства каркасных белков Cas, был выбран для дальнейшего исследования, поскольку было показано, что он участвует в подвижности NC 24 , хотя механизм действия неизвестен. Коиммунопреципитация подтвердила ассоциацию эндогенных белков DLC1 и NEDD9 в эмбриональных нервных трубках (рис. 5d), что повышает вероятность того, что DN-DLC1 может мешать взаимодействию DLC1-NEDD9, исходя из способа его действия из предыдущего исследования 21 . В самом деле, эктопическая экспрессия DN-DLC1 также способна коиммунопреципитировать с NEDD9.Дальнейшее картирование доменов показало, что усеченная на С-конце версия DN-DLC1, состоящая из доменов SAM и FAT [DN-DLC1 (320)] 25 , где большинство партнеров по взаимодействию связывания DLC1 могут связываться с NEDD9, тогда как домен SAM сам по себе не показать взаимодействие с NEDD9 (рис. 5e). Соответственно, сверхэкспрессия DN-DLC1 или DN-DLC1 (320), но не только домена SAM, способна снижать отслаивание NC (Supplementary Fig. 6a). Эти результаты предполагают, что NEDD9 может быть изолирован от его эндогенного взаимодействия с DLC1 посредством связывания с доменом FAT DN-DLC1.
5e). Соответственно, сверхэкспрессия DN-DLC1 или DN-DLC1 (320), но не только домена SAM, способна снижать отслаивание NC (Supplementary Fig. 6a). Эти результаты предполагают, что NEDD9 может быть изолирован от его эндогенного взаимодействия с DLC1 посредством связывания с доменом FAT DN-DLC1.
NEDD9 как белок, взаимодействующий с DLC1, необходим для поляризованной экспрессии DLC1. — диаграмма Венна , показывающая количество белков, обогащенных DLC1. b DLC1-ассоциированные белки, участвующие в основных канонических путях, согласно оценке базы знаний Ingenuity Pathway Knowledge. c Интерактомная сеть DLC1 и связанных с ней белков, участвующих в клеточной подвижности и регуляции цитоскелета. d Иммунопреципитацию (IP) проводили в лизате нервной трубки с использованием анти-DLC1 и IgG в качестве контроля.Иммунопреципитированные белки разделяли на гелях для электрофореза в полиакриламидном геле с додецилсульфатом натрия и проводили вестерн-блоттинг с антителами против NEDD9 и DLC1. e Схематическое изображение конструкции усеченных с С-конца конструкций DN-DLC1. IP с анти-V5 проводили в лизатах, экстрагированных из нервных трубок, электропорированных с указанными конструкциями, с последующим вестерн-блоттингом с анти-V5 или анти-NEDD9. Белая стрелка указывает на полосу специфического белка DN-DLC1 (SAM). f Гибридизация in situ для NEDD9 , DLC1 , SOX9 и SOX10 на поперечных срезах куриного эмбриона st 11. Иммунофлуоресценция для NEDD9, SOX9 и SOX10 на поперечных срезах куриного эмбриона st 11. Масштабная линейка: 50 мкм. г Иммунофлуоресценция для NEDD9 и фаллоидина на эмигрирующих NCC из эксплантатов нервной трубки и ядра окрашивают DAPI. Поляризованная экспрессия NEDD9 показана псевдоцветом. Анализ линейного сканирования, показывающий среднюю интенсивность флуоресценции NEDD9 вдоль белой пунктирной линии ( n = 103/11 эксплантов). ч Иммунофлуоресценция для DLC1 и фаллоидина на NCC, экспрессирующих контрольный морфолино (Ctrl-MO) ( n = 65/11 эксплантатов) или NEDD9-MO ( n = 112/12 эксплантов).
e Схематическое изображение конструкции усеченных с С-конца конструкций DN-DLC1. IP с анти-V5 проводили в лизатах, экстрагированных из нервных трубок, электропорированных с указанными конструкциями, с последующим вестерн-блоттингом с анти-V5 или анти-NEDD9. Белая стрелка указывает на полосу специфического белка DN-DLC1 (SAM). f Гибридизация in situ для NEDD9 , DLC1 , SOX9 и SOX10 на поперечных срезах куриного эмбриона st 11. Иммунофлуоресценция для NEDD9, SOX9 и SOX10 на поперечных срезах куриного эмбриона st 11. Масштабная линейка: 50 мкм. г Иммунофлуоресценция для NEDD9 и фаллоидина на эмигрирующих NCC из эксплантатов нервной трубки и ядра окрашивают DAPI. Поляризованная экспрессия NEDD9 показана псевдоцветом. Анализ линейного сканирования, показывающий среднюю интенсивность флуоресценции NEDD9 вдоль белой пунктирной линии ( n = 103/11 эксплантов). ч Иммунофлуоресценция для DLC1 и фаллоидина на NCC, экспрессирующих контрольный морфолино (Ctrl-MO) ( n = 65/11 эксплантатов) или NEDD9-MO ( n = 112/12 эксплантов). Распределение экспрессии DLC1 в обоих вариантах лечения показано псевдоцветом. Масштабные линейки 20 мкм. i Анализ линейного сканирования, показывающий среднюю интенсивность флуоресценции DLC1 в обработанных NEDD9-MO NCC ( n = 112/12 эксплантов)
Распределение экспрессии DLC1 в обоих вариантах лечения показано псевдоцветом. Масштабные линейки 20 мкм. i Анализ линейного сканирования, показывающий среднюю интенсивность флуоресценции DLC1 в обработанных NEDD9-MO NCC ( n = 112/12 эксплантов)
Гибридизация in situ на последовательных срезах показывает, что мРНК NEDD9 была обнаружена в предмиграционных и ранних мигрирующих стволовых NCC эмбрионов st 11 аналогично DLC1 , SOX9 и SOX10 .Кроме того, выражение NEDD9 также было заметно в пластине пола. Распределение транскриптов NEDD9 напоминает его экспрессию белка, которая была обнаружена в цитоплазме пред мигрирующих и эмигрирующих NCC, перекрывая ядерную локализацию белков SOX9 и SOX10, что указывает на совместную локализацию этих факторов в начале отслоения NC Рис. 5е). Иммунофлуоресцентный анализ показал, что NEDD9 демонстрирует картину, аналогичную DLC1, с поляризованной локализацией между передним краем и ядром эмигрирующих NCC (рис. 5г). Затем мы исследовали, изменяется ли пространственная экспрессия DLC1 в клетках, обработанных морфолино NEDD9 (NEDD9-MO), что снижает уровни белка NEDD9 в расслаивающихся NCC, по сравнению с нетрансфицированной стороной и эмбрионами, обработанными контрольным-MO (Ctrl-MO) (Дополнительный рис. 7а). NEDD9-MO приводит к аберрантной и удлиненной морфологии NCC без заметной полярности спереди назад (рис. 5h). Важно отметить, что белок DLC1 демонстрировал случайное распределение в клетках, трансфицированных NEDD9-MO, как показывает анализ линейного сканирования, в то время как экспрессия поляризованного DLC1 оставалась неизменной в клетках, обработанных Ctrl-MO (рис.5h, i), предполагая, что асимметричная локализация DLC1 регулируется ассоциацией с его партнером по связыванию NEDD9.
5г). Затем мы исследовали, изменяется ли пространственная экспрессия DLC1 в клетках, обработанных морфолино NEDD9 (NEDD9-MO), что снижает уровни белка NEDD9 в расслаивающихся NCC, по сравнению с нетрансфицированной стороной и эмбрионами, обработанными контрольным-MO (Ctrl-MO) (Дополнительный рис. 7а). NEDD9-MO приводит к аберрантной и удлиненной морфологии NCC без заметной полярности спереди назад (рис. 5h). Важно отметить, что белок DLC1 демонстрировал случайное распределение в клетках, трансфицированных NEDD9-MO, как показывает анализ линейного сканирования, в то время как экспрессия поляризованного DLC1 оставалась неизменной в клетках, обработанных Ctrl-MO (рис.5h, i), предполагая, что асимметричная локализация DLC1 регулируется ассоциацией с его партнером по связыванию NEDD9.
Ассоциация DLC1 с NEDD9 ограничивает активный RhoA
Поскольку NEDD9 участвует в регуляции Rho GTPases в раковых клетках 26 , вышеуказанные открытия открывают две возможности. Утрата функции NEDD9 может приводить к аберрантной активности RHOA, что приводит к потере полярности клеток. С другой стороны, неправильная локализация DLC1 является основной причиной дисрегуляции полярности NCC в NEDD9-MO.Чтобы различить эти возможности, мы сначала исследовали активность RHOA в отслаивании NCCs, экспрессирующих NEDD9-MO in vivo, и в эксплантатах нервной трубки, обработанных SDF-1 (Fig. 6a, c). Неожиданно количественная оценка общей активности FRET в клетках, обработанных NEDD9-MO in vivo, была сопоставима с векторным контролем и клетками со сверхэкспрессией NEDD9 (рис. 6b), что позволяет предположить, что потеря функции NEDD9 и повышенный уровень экспрессии NEDD9 не имеют прямого влияния на общую Деятельность RHOA. Однако мы наблюдали неправильную локализацию активности RHOA вокруг ядра клетки, обработанной NEDD9-MO, которая демонстрировала аберрантную поляризацию спереди назад и отсутствие направленности в миграции (рис.6a, c и дополнительный фильм 10). Соответственно, индекс FRET между задней и передней частью обработанных NEDD9-MO клеток не проявлял дифференциального характера по сравнению с контролем и клетками, сверхэкспрессирующими NEDD9 (Fig.
С другой стороны, неправильная локализация DLC1 является основной причиной дисрегуляции полярности NCC в NEDD9-MO.Чтобы различить эти возможности, мы сначала исследовали активность RHOA в отслаивании NCCs, экспрессирующих NEDD9-MO in vivo, и в эксплантатах нервной трубки, обработанных SDF-1 (Fig. 6a, c). Неожиданно количественная оценка общей активности FRET в клетках, обработанных NEDD9-MO in vivo, была сопоставима с векторным контролем и клетками со сверхэкспрессией NEDD9 (рис. 6b), что позволяет предположить, что потеря функции NEDD9 и повышенный уровень экспрессии NEDD9 не имеют прямого влияния на общую Деятельность RHOA. Однако мы наблюдали неправильную локализацию активности RHOA вокруг ядра клетки, обработанной NEDD9-MO, которая демонстрировала аберрантную поляризацию спереди назад и отсутствие направленности в миграции (рис.6a, c и дополнительный фильм 10). Соответственно, индекс FRET между задней и передней частью обработанных NEDD9-MO клеток не проявлял дифференциального характера по сравнению с контролем и клетками, сверхэкспрессирующими NEDD9 (Fig. 6b-d, f). Эти данные предполагают, что отсутствие полярности RHOA в отсутствие функции NEDD9 может быть связано с нерегулируемой локализацией DLC1, которая нарушает точное пространственное ограничение RHOA без изменения общего уровня активности RHOA. Это побудило нас дополнительно изучить, является ли ассоциация DLC1 с NEDD9 функционально необходимой для установления полярности RHOA.Затем мы сверхэкспрессировали DN-DLC1, чтобы нарушить взаимодействие эндогенного DLC1 с NEDD9, делая DLC1 неспособным к пространственному ограничению активности RHOA в задней части клетки. Если бы это было так, то поляризованная активность RHOA могла бы быть восстановлена сверхэкспрессией NEDD9 в экспрессирующих DN-DLC1 клетках. Действительно, дифференциальное распределение активности RHOA между задней и передней частью в клетках, экспрессирующих DN-DLC1 + NEDD9 in vivo, было восстановлено с индексом FRET, сравнимым с контролем, и только сверхэкспрессией NEDD9, но его общий сигнал FRET оставался выше, чем при других обработках (рис.
6b-d, f). Эти данные предполагают, что отсутствие полярности RHOA в отсутствие функции NEDD9 может быть связано с нерегулируемой локализацией DLC1, которая нарушает точное пространственное ограничение RHOA без изменения общего уровня активности RHOA. Это побудило нас дополнительно изучить, является ли ассоциация DLC1 с NEDD9 функционально необходимой для установления полярности RHOA.Затем мы сверхэкспрессировали DN-DLC1, чтобы нарушить взаимодействие эндогенного DLC1 с NEDD9, делая DLC1 неспособным к пространственному ограничению активности RHOA в задней части клетки. Если бы это было так, то поляризованная активность RHOA могла бы быть восстановлена сверхэкспрессией NEDD9 в экспрессирующих DN-DLC1 клетках. Действительно, дифференциальное распределение активности RHOA между задней и передней частью в клетках, экспрессирующих DN-DLC1 + NEDD9 in vivo, было восстановлено с индексом FRET, сравнимым с контролем, и только сверхэкспрессией NEDD9, но его общий сигнал FRET оставался выше, чем при других обработках (рис. .6а, б). Кластеризация индекса FRET между задней и передней частью культивируемых NCC, экспрессирующих DN-DLC1 + NEDD9, также показала восстановление дифференциальной активности RHOA, такой как клетки, сверхэкспрессирующие NEDD9, и клетки продемонстрировали поляризованную морфологию и направленную миграцию (рис. 6c, e, f и дополнительные фильмы. 11 и 12). В соответствии с этим, восстановление расслоения NC наблюдалось у эмбрионов, электропорированных с помощью DN-DLC1 + NEDD9, в отличие от заметного уменьшения SOX10 Клетки + и HNK-1 + на трансфицированной стороне эмбрионов, подвергнутых электропорации с помощью DN-DLC1 или NEDD9-MO, тогда как сверхэкспрессия только NEDD9 не влияла на отслоение NC (рис.6г, з). В совокупности эти данные показывают, что взаимодействие DLC1 с NEDD9 необходимо для пространственного ограничения активности RHOA в задней части клетки и на переднем крае, что является предпосылкой для установления оси полярности спереди-сзади и направленного отслоения и миграции ствола.
.6а, б). Кластеризация индекса FRET между задней и передней частью культивируемых NCC, экспрессирующих DN-DLC1 + NEDD9, также показала восстановление дифференциальной активности RHOA, такой как клетки, сверхэкспрессирующие NEDD9, и клетки продемонстрировали поляризованную морфологию и направленную миграцию (рис. 6c, e, f и дополнительные фильмы. 11 и 12). В соответствии с этим, восстановление расслоения NC наблюдалось у эмбрионов, электропорированных с помощью DN-DLC1 + NEDD9, в отличие от заметного уменьшения SOX10 Клетки + и HNK-1 + на трансфицированной стороне эмбрионов, подвергнутых электропорации с помощью DN-DLC1 или NEDD9-MO, тогда как сверхэкспрессия только NEDD9 не влияла на отслоение NC (рис.6г, з). В совокупности эти данные показывают, что взаимодействие DLC1 с NEDD9 необходимо для пространственного ограничения активности RHOA в задней части клетки и на переднем крае, что является предпосылкой для установления оси полярности спереди-сзади и направленного отслоения и миграции ствола. НКЦ.
НКЦ.
Взаимодействие между NEDD9 и DLC1 необходимо для полярности RHOA и направленной миграции NC. a Обработанные изображения сигналов FRET поперечных сечений эмбрионов, электропорированных с помощью датчика FRET плюс NEDD9-MO ( n = 146/17 эмбрионов), NEDD9 ( n = 168/13 эмбрионов) или DN-DLC1 + NEDD9 ( n = 152/13 эмбрионов).NCC маркируются иммунофлуоресценцией SOX9, а ядра окрашиваются DAPI. Увеличенная область и выбранные ячейки отмечены пунктирными квадратами. Белые стрелки указывают на сигнал FRET. Масштабные линейки: 20 мкм. b Количественная оценка общего индекса FRET NCC, подвергнутых электропорации с указанными конструкциями. Количественная оценка отношения индекса FRET между задней и передней частью NCC, электропорированных с указанными конструкциями (контроль, n = 425/25 эмбрионов). Среднее ± s.e.m .; Студенческий t -тест, ** p <0.001; Тест множественного сравнения Бонферрони, *** p <0,0001 c Обработанные изображения сигнала FRET покадровой серии мигрирующих NCC, экспрессирующих NEDD9-MO ( n = 97/15 эксплантатов), DN-DLC1 + NEDD9-MO ( n = 92/19 эксплантов) и NEDD9 ( n = 106/16 эксплантов). Масштабные линейки: 20 мкм. d Тепловые карты, представляющие индекс FRET как функцию времени на задней или передней панели, и соотношение FRET между задней и передней частью 12 выбранных NCC, выражающих NEDD9-MO, e DN-DLC1 + NEDD9 и f NEDD9. г Гибридизация in situ SOX10 3’UTR в эмбрионах, электропорированных с DN-DLC1 ( n = 9), NEDD9 ( n = 10), NEDD9-MO ( n = 10) и DN- DLC1 + NEDD9 ( n = 9). Черные стрелки указывают на снижение экспрессии SOX103’UTR на трансфицированных сторонах. Масштабная линейка 10 мкм. ч Количественная оценка количества GFP + NCC, экспрессирующих HNK-1 + , в эмбрионах, подвергнутых электропорации с указанными конструкциями.Среднее ± s.e.m. Тест множественного сравнения Бонферрони, ** p <0,001; *** р <0,0001
Масштабные линейки: 20 мкм. d Тепловые карты, представляющие индекс FRET как функцию времени на задней или передней панели, и соотношение FRET между задней и передней частью 12 выбранных NCC, выражающих NEDD9-MO, e DN-DLC1 + NEDD9 и f NEDD9. г Гибридизация in situ SOX10 3’UTR в эмбрионах, электропорированных с DN-DLC1 ( n = 9), NEDD9 ( n = 10), NEDD9-MO ( n = 10) и DN- DLC1 + NEDD9 ( n = 9). Черные стрелки указывают на снижение экспрессии SOX103’UTR на трансфицированных сторонах. Масштабная линейка 10 мкм. ч Количественная оценка количества GFP + NCC, экспрессирующих HNK-1 + , в эмбрионах, подвергнутых электропорации с указанными конструкциями.Среднее ± s.e.m. Тест множественного сравнения Бонферрони, ** p <0,001; *** р <0,0001
SOXE регулируют экспрессию генов DLC1 и NEDD9
Факторы транскрипции SOXE SOX9 и SOX10 играют важную роль в регуляции спецификации NC ствола и отслаивания 27, 28 . Перекрывающаяся экспрессия DLC1 , NEDD9 , SOX9 и SOX10 в премиграционных и ранних мигрирующих NCC побудила нас исследовать, подвергаются ли экспрессии DLC1 и NEDD9 транскрипции / регуляции SOX9 и SOX10.Результаты показали, что SOX9 или SOX10 были достаточными для индукции эктопической экспрессии DLC1 по всей нервной трубке на 12-24 hpt, но не на 6 hpt (Fig. 7a and Supplementary Fig. 8a, b). Принимая во внимание, что SOX9 индуцировал экспрессию SOX10 29 , DLC1 не индуцировался в нервной трубке, подвергнутой электропорации с SOX9 плюс SOX10-MO через 24 ч (дополнительный рисунок 8c). Соответственно, подавление экспрессии DLC1 наблюдалось в эмбрионах, обработанных SOX10-MO, по сравнению с Ctrl-MO (рис.7б). Эти результаты показывают, что SOX9 индуцирует SOX10 для регулирования экспрессии DLC1 . Для дальнейшего исследования, может ли DLC1 опосредовать SOX9 и SOX10-индуцированное расслоение и миграцию NC, мы электропорировали SOX9 или SOX10 плюс DN-DLC1 и исследовали их влияние на подвижность NC, используя FOXD3 в качестве мигрирующего маркера NC (рис.
Перекрывающаяся экспрессия DLC1 , NEDD9 , SOX9 и SOX10 в премиграционных и ранних мигрирующих NCC побудила нас исследовать, подвергаются ли экспрессии DLC1 и NEDD9 транскрипции / регуляции SOX9 и SOX10.Результаты показали, что SOX9 или SOX10 были достаточными для индукции эктопической экспрессии DLC1 по всей нервной трубке на 12-24 hpt, но не на 6 hpt (Fig. 7a and Supplementary Fig. 8a, b). Принимая во внимание, что SOX9 индуцировал экспрессию SOX10 29 , DLC1 не индуцировался в нервной трубке, подвергнутой электропорации с SOX9 плюс SOX10-MO через 24 ч (дополнительный рисунок 8c). Соответственно, подавление экспрессии DLC1 наблюдалось в эмбрионах, обработанных SOX10-MO, по сравнению с Ctrl-MO (рис.7б). Эти результаты показывают, что SOX9 индуцирует SOX10 для регулирования экспрессии DLC1 . Для дальнейшего исследования, может ли DLC1 опосредовать SOX9 и SOX10-индуцированное расслоение и миграцию NC, мы электропорировали SOX9 или SOX10 плюс DN-DLC1 и исследовали их влияние на подвижность NC, используя FOXD3 в качестве мигрирующего маркера NC (рис. 7c, d). В то время как сверхэкспрессия SOX9 или SOX10 способствовала надежному отслаиванию и миграции NC, электропорация SOX9 + DN-DLC1 привела к почти полному устранению отслоения и миграции NC (рис.7в, г). Эффект был относительно слабым у эмбрионов, электропорированных с SOX10 + DN-DLC1 с уменьшенным FOXD3 + мигрирующих NCC по сравнению с нетрансфицированной стороной (рис. 7d). Вместе эти данные демонстрируют, что SOX10 способствует миграции NC посредством регулирования DLC1 . Затем мы исследовали, регулируют ли SOX9 и / или SOX10 экспрессию NEDD9 . Электропорация SOX9 привела к эктопической экспрессии NEDD9 на 6 и 12 hpt по всей дорсально-вентральной оси нервной трубки по сравнению с неэлектропорированной стороной (рис.7e и дополнительный рис. 8d). Кроме того, клетки, экспрессирующие SOX9, были совместно локализованы с эктопической экспрессией белка NEDD9, что указывает на то, что SOX9 индуцирует экспрессию NEDD9 клеточно-автономным образом (дополнительный рис.
7c, d). В то время как сверхэкспрессия SOX9 или SOX10 способствовала надежному отслаиванию и миграции NC, электропорация SOX9 + DN-DLC1 привела к почти полному устранению отслоения и миграции NC (рис.7в, г). Эффект был относительно слабым у эмбрионов, электропорированных с SOX10 + DN-DLC1 с уменьшенным FOXD3 + мигрирующих NCC по сравнению с нетрансфицированной стороной (рис. 7d). Вместе эти данные демонстрируют, что SOX10 способствует миграции NC посредством регулирования DLC1 . Затем мы исследовали, регулируют ли SOX9 и / или SOX10 экспрессию NEDD9 . Электропорация SOX9 привела к эктопической экспрессии NEDD9 на 6 и 12 hpt по всей дорсально-вентральной оси нервной трубки по сравнению с неэлектропорированной стороной (рис.7e и дополнительный рис. 8d). Кроме того, клетки, экспрессирующие SOX9, были совместно локализованы с эктопической экспрессией белка NEDD9, что указывает на то, что SOX9 индуцирует экспрессию NEDD9 клеточно-автономным образом (дополнительный рис. 8d). В соответствии с этим истощение экспрессии SOX9 в обработанных SOX9-MO эмбрионах привело к подавлению экспрессии NEDD9 за 24 часа по сравнению с Ctrl-MO (рис. 7f), указывая на то, что SOX9 необходим для экспрессии NEDD9 . Напротив, истощение экспрессии белка SOX10 под действием Sox10-MO не повлияло на уровни экспрессии SOX9 и NEDD9 , а экспрессия NEDD9 все еще индуцировалась в клетках, экспрессирующих SOX9 + SOX10-MO (дополнительный рис.8e, f), предполагая, что SOX10 не требуется для экспрессии NEDD9 . Соответственно, сверхэкспрессия SOX10 не способна индуцировать эктопическую экспрессию NEDD9, хотя она запускает экспрессию HNK-1 через 12 hpt (дополнительный рис. 8g). Иммунопреципитация хроматина и репортерные анализы показали, что NEDD9 является прямой транскрипционной мишенью SOX9, но не SOX10 посредством связывания с последовательностями энхансера NEDD9 , но не с промоторной областью (Рис. 7g, h и Дополнительный Рис. 9a – d).
8d). В соответствии с этим истощение экспрессии SOX9 в обработанных SOX9-MO эмбрионах привело к подавлению экспрессии NEDD9 за 24 часа по сравнению с Ctrl-MO (рис. 7f), указывая на то, что SOX9 необходим для экспрессии NEDD9 . Напротив, истощение экспрессии белка SOX10 под действием Sox10-MO не повлияло на уровни экспрессии SOX9 и NEDD9 , а экспрессия NEDD9 все еще индуцировалась в клетках, экспрессирующих SOX9 + SOX10-MO (дополнительный рис.8e, f), предполагая, что SOX10 не требуется для экспрессии NEDD9 . Соответственно, сверхэкспрессия SOX10 не способна индуцировать эктопическую экспрессию NEDD9, хотя она запускает экспрессию HNK-1 через 12 hpt (дополнительный рис. 8g). Иммунопреципитация хроматина и репортерные анализы показали, что NEDD9 является прямой транскрипционной мишенью SOX9, но не SOX10 посредством связывания с последовательностями энхансера NEDD9 , но не с промоторной областью (Рис. 7g, h и Дополнительный Рис. 9a – d). Таким образом, эти результаты указывают на специфическую роль SOX9 в прямой регуляции транскрипции экспрессии NEDD9 . Соответственно, анализ эпистаза показал, что клетки, экспрессирующие SOX9 + NEDD9-MO, оставались в нейроэпителии, а количество мигрирующих NCC HNK-1 + на трансфицированной стороне было снижено по сравнению с векторным контролем и эмбрионы, электропорированные с SOX9 или SOX9 + Ctrl- МО (рис. 7ж, к). Взятые вместе, эти результаты показывают, что SOX9 способствует отслаиванию и миграции NC посредством прямой транскрипционной активации экспрессии NEDD9 .
Таким образом, эти результаты указывают на специфическую роль SOX9 в прямой регуляции транскрипции экспрессии NEDD9 . Соответственно, анализ эпистаза показал, что клетки, экспрессирующие SOX9 + NEDD9-MO, оставались в нейроэпителии, а количество мигрирующих NCC HNK-1 + на трансфицированной стороне было снижено по сравнению с векторным контролем и эмбрионы, электропорированные с SOX9 или SOX9 + Ctrl- МО (рис. 7ж, к). Взятые вместе, эти результаты показывают, что SOX9 способствует отслаиванию и миграции NC посредством прямой транскрипционной активации экспрессии NEDD9 .
Факторы SOXE регулируют экспрессию DLC1 и NEDD9 . a Гибридизация in situ для DLC1 на поперечных срезах эмбрионов, электропорированных с SOX10 на 12 hpt ( n = 9). Масштабная линейка, 50 мкм. b Гибридизация in situ для DLC1 на эмбрионах, электропорированных с помощью Ctrl-MO ( n = 10) или SOX10-MO ( n = 10) на 24hpt. Черные стрелки указывают на потерю экспрессии DLC1 на трансфицированной стороне нервной трубки.Масштабная линейка 10 мкм. c Гибридизация in situ для FOXD3 на эмбрионах, электропорированных с SOX9 ( n = 9) или SOX9 + DN-DLC1 ( n = 9) на 24hpt. Масштабная линейка, 20 мкм d Гибридизация in situ для FOXD3 на эмбрионах, электропорированных с SOX10 ( n = 10) или SOX10 + DN-DLC1 ( n = 10) при 24hpt. Масштабная линейка 10 мкм. e Иммунофлуоресценция для NEDD9 и гибридизация in situ для NEDD9 на поперечных срезах эмбрионов, электропорированных с SOX9 ( n = 12) на 6 hpt.Масштабная линейка, 50 мкм. f Иммунофлуоресценция для SOX9 и гибридизация in situ для NEDD9 на поперечных срезах эмбрионов, электропорированных с помощью Ctrl-MO ( n = 9) или SOX9-MO ( n = 9). Белая стрелка указывает на потерю экспрессии SOX9 на трансфицированной стороне. Масштабная линейка: 50 мкм.
Черные стрелки указывают на потерю экспрессии DLC1 на трансфицированной стороне нервной трубки.Масштабная линейка 10 мкм. c Гибридизация in situ для FOXD3 на эмбрионах, электропорированных с SOX9 ( n = 9) или SOX9 + DN-DLC1 ( n = 9) на 24hpt. Масштабная линейка, 20 мкм d Гибридизация in situ для FOXD3 на эмбрионах, электропорированных с SOX10 ( n = 10) или SOX10 + DN-DLC1 ( n = 10) при 24hpt. Масштабная линейка 10 мкм. e Иммунофлуоресценция для NEDD9 и гибридизация in situ для NEDD9 на поперечных срезах эмбрионов, электропорированных с SOX9 ( n = 12) на 6 hpt.Масштабная линейка, 50 мкм. f Иммунофлуоресценция для SOX9 и гибридизация in situ для NEDD9 на поперечных срезах эмбрионов, электропорированных с помощью Ctrl-MO ( n = 9) или SOX9-MO ( n = 9). Белая стрелка указывает на потерю экспрессии SOX9 на трансфицированной стороне. Масштабная линейка: 50 мкм. g Схематическая диаграмма, показывающая семь экзонов ( черный ) гена цыпленка NEDD9 с промоторной областью синего цвета, энхансер 1 -9,5 т.п.н. энхансер 2 (E2), содержащий предполагаемый связывающий мотив SOX9 , B3. ч Кратное обогащение трех независимых анализов ChIP-qPCR на SOX9, SOX10 и IgG (контроль) антител к B1, B2 и B3. Данные представлены в виде кратного обогащения для предполагаемых SOX9 связывающих мотивов относительно контроля. Среднее ± s.e.m. Тест множественного сравнения Бонферрони, * p <0,05, ** p <0,001, *** p <0,0001. i Кратковременная активация трех независимых анализов люциферазы in vivo для репортерных конструкций, несущих NEDD9 -промотор, E1 или E2.Среднее ± s.e.m. Тест множественного сравнения Бонферрони, ** p <0,001, *** p <0,0001. j Иммунофлуоресценция для HNK-1, ламинина и гибридизация in situ для SOX103’UTR на поперечных срезах эмбрионов, электропорированных с SOX9 или SOX9 + NEDD9-MO при 24hpt.
g Схематическая диаграмма, показывающая семь экзонов ( черный ) гена цыпленка NEDD9 с промоторной областью синего цвета, энхансер 1 -9,5 т.п.н. энхансер 2 (E2), содержащий предполагаемый связывающий мотив SOX9 , B3. ч Кратное обогащение трех независимых анализов ChIP-qPCR на SOX9, SOX10 и IgG (контроль) антител к B1, B2 и B3. Данные представлены в виде кратного обогащения для предполагаемых SOX9 связывающих мотивов относительно контроля. Среднее ± s.e.m. Тест множественного сравнения Бонферрони, * p <0,05, ** p <0,001, *** p <0,0001. i Кратковременная активация трех независимых анализов люциферазы in vivo для репортерных конструкций, несущих NEDD9 -промотор, E1 или E2.Среднее ± s.e.m. Тест множественного сравнения Бонферрони, ** p <0,001, *** p <0,0001. j Иммунофлуоресценция для HNK-1, ламинина и гибридизация in situ для SOX103’UTR на поперечных срезах эмбрионов, электропорированных с SOX9 или SOX9 + NEDD9-MO при 24hpt. Белая стрелка указывает на потерю экспрессии ламинина на базальной мембране трансфицированной нервной трубки. Масштабная линейка: 50 мкм. k Количественная оценка количества эмигрирующих GFP + NCC, экспрессирующих HNK-1, в эмбрионах, электропорированных с указанными конструкциями.Среднее ± s.e.m. Тест множественного сравнения Бонферрони, * p <0,05; ** p <0,001; нс, незначительно
Белая стрелка указывает на потерю экспрессии ламинина на базальной мембране трансфицированной нервной трубки. Масштабная линейка: 50 мкм. k Количественная оценка количества эмигрирующих GFP + NCC, экспрессирующих HNK-1, в эмбрионах, электропорированных с указанными конструкциями.Среднее ± s.e.m. Тест множественного сравнения Бонферрони, * p <0,05; ** p <0,001; нс, незначительно
FRET с использованием активирующих сперматозоидов пептидов, меченных флуоресцентными белками, показывает, что лиганд-связывающие сайты существуют в виде кластеров.
РЕЗЮМЕ
Клеточная связь на большом расстоянии между спермой и яйцеклеткой имеет решающее значение для внешнего оплодотворения. Пептиды, активирующие сперматозоиды (SAP), представляют собой диффундирующие компоненты внешнего слоя яиц у иглокожих и действуют как хемоаттрактанты для сперматозоидов.Декапептид, названный speract, является наиболее охарактеризованным SAP морского ежа. Биохимические и физиологические действия speract были изучены с очищенными или химически синтезированными пептидами. В этой работе мы приготовили рекомбинантный спектр, слитый с флуоресцентным белком (FP; FP-speract), с использованием трех цветовых вариантов: голубого (eCFP), желтого (mVenus) и большого желтого стоксова сдвига (mAmetrine) FP. Хотя эти флуоресцентные метки в 20 раз больше, чем speract, эксперименты по конкурентному связыванию с использованием mAmetrine-speract показали, что этот FP-speract имеет сопоставимое сродство связывания с рецептором (7.В 6 раз меньше) по сравнению с немеченым speract. Действительно, 10 нмоль -1 eCFP-speract индуцирует физиологические реакции сперматозоидов, такие как изменения мембранного потенциала и повышение внутриклеточного pH и концентрации Ca 2+ , аналогичные тем, которые вызываются 10 нмоль l -1 speract. Кроме того, FP-speract сохраняет свою флуоресценцию после связывания со своим рецептором. Используя это свойство, мы провели измерения флуоресцентного резонансного переноса энергии (FRET) с eCFP-speract и mVenus-speract в качестве зондов и получили положительный сигнал FRET при связывании с рецептором, что позволяет предположить, что рецептор speract существует в виде олигомера, по крайней мере, как димер, или, альтернативно, что один рецепторный белок speract обладает множеством сайтов связывания.
Биохимические и физиологические действия speract были изучены с очищенными или химически синтезированными пептидами. В этой работе мы приготовили рекомбинантный спектр, слитый с флуоресцентным белком (FP; FP-speract), с использованием трех цветовых вариантов: голубого (eCFP), желтого (mVenus) и большого желтого стоксова сдвига (mAmetrine) FP. Хотя эти флуоресцентные метки в 20 раз больше, чем speract, эксперименты по конкурентному связыванию с использованием mAmetrine-speract показали, что этот FP-speract имеет сопоставимое сродство связывания с рецептором (7.В 6 раз меньше) по сравнению с немеченым speract. Действительно, 10 нмоль -1 eCFP-speract индуцирует физиологические реакции сперматозоидов, такие как изменения мембранного потенциала и повышение внутриклеточного pH и концентрации Ca 2+ , аналогичные тем, которые вызываются 10 нмоль l -1 speract. Кроме того, FP-speract сохраняет свою флуоресценцию после связывания со своим рецептором. Используя это свойство, мы провели измерения флуоресцентного резонансного переноса энергии (FRET) с eCFP-speract и mVenus-speract в качестве зондов и получили положительный сигнал FRET при связывании с рецептором, что позволяет предположить, что рецептор speract существует в виде олигомера, по крайней мере, как димер, или, альтернативно, что один рецепторный белок speract обладает множеством сайтов связывания. Это свойство может частично объяснять положительное и / или отрицательное совместное связывание speract с рецептором.
Это свойство может частично объяснять положительное и / или отрицательное совместное связывание speract с рецептором.
ВВЕДЕНИЕ
У большинства морских животных сперматозоиды и яйца выбрасываются в океан, где расстояние между гаметами увеличивается при разбавлении, что подразумевает потенциальную проблему в достижении успешного оплодотворения для любого вида. Чтобы решить эту проблему, морские животные обычно производят большое количество гамет, хотя существует компромисс между их количеством и размером.В случае иглокожих женские гаметы используют две дополнительные стратегии для улучшения взаимодействий сперматозоидов. Один из них — увеличение физических размеров за счет набухания внеклеточного матрикса, называемого яичным желе, за счет поглощения морской воды (Подольский, 2002). Другой — высвобождение специфических хемоаттрактантов для сперматозоидов, пептидов, активирующих сперматозоиды (SAP) (Suzuki, 1995). SAP инициируют свое физиологическое действие на сперматозоиды за счет повышения уровня циклического гуанозинмонофосфата (цГМФ) (Garbers, 1989; Kaupp et al. , 2003), который вызывает изменения мембранного потенциала ( V m ) (Galindo et al., 2000; Strünker et al., 2006), что приводит к увеличению внутриклеточного pH (pH i ) и Ca 2+. концентраций ([Ca 2+ ] i ) (Nishigaki et al., 2001; Seifert et al., 2015). Изменения в [Ca 2+ ] и , в частности, играют фундаментальную роль в регуляции формы биения жгутика (Böhmer et al., 2005; Wood et al., 2005), а именно в процессе образования сперматозоидов. хемотаксис (Alvarez et al., 2012; Guerrero et al., 2010).
, 2003), который вызывает изменения мембранного потенциала ( V m ) (Galindo et al., 2000; Strünker et al., 2006), что приводит к увеличению внутриклеточного pH (pH i ) и Ca 2+. концентраций ([Ca 2+ ] i ) (Nishigaki et al., 2001; Seifert et al., 2015). Изменения в [Ca 2+ ] и , в частности, играют фундаментальную роль в регуляции формы биения жгутика (Böhmer et al., 2005; Wood et al., 2005), а именно в процессе образования сперматозоидов. хемотаксис (Alvarez et al., 2012; Guerrero et al., 2010).
С тех пор, как первый SAP (speract) был очищен и структурно определен (Suzuki, 1995), было идентифицировано почти 100 SAP у многих видов иглокожих (Nishigaki et al., 1996; Suzuki, 1995). SAP делятся на пять или шесть групп в зависимости от их структуры и перекрестной деятельности (Suzuki, 1995). Существует много изоформ в одном и том же классе SAP, учитывая существование изоформ внутри и между видами. Разнообразие SAP внутри видов связано с биосинтезом этих пептидов.МРНК, кодирующая speract и asterosap (SAP, выделенная из морской звезды Asterias amurensis ), показала, что SAP продуцируются как большие белки-предшественники, содержащие множество изоформ SAP: 9–12 изоформ speract, разделенных одной аминокислотой (Lys) (Kinoh et al. ., 1994; Ramarao et al., 1990) и приблизительно 10 изоформ asterosap, разделенных спейсерным пептидом из 12–22 аминокислот (Matsumoto et al., 1999; Nakachi et al., 2008). Также сообщалось о множественных копиях одного и того же пептида в одном белке-предшественнике для сарафотоксина в яде змеи (Ducancel et al., 1993), предлагая эффективную стратегию получения высокой концентрации подобных пептидов.
Разнообразие SAP внутри видов связано с биосинтезом этих пептидов.МРНК, кодирующая speract и asterosap (SAP, выделенная из морской звезды Asterias amurensis ), показала, что SAP продуцируются как большие белки-предшественники, содержащие множество изоформ SAP: 9–12 изоформ speract, разделенных одной аминокислотой (Lys) (Kinoh et al. ., 1994; Ramarao et al., 1990) и приблизительно 10 изоформ asterosap, разделенных спейсерным пептидом из 12–22 аминокислот (Matsumoto et al., 1999; Nakachi et al., 2008). Также сообщалось о множественных копиях одного и того же пептида в одном белке-предшественнике для сарафотоксина в яде змеи (Ducancel et al., 1993), предлагая эффективную стратегию получения высокой концентрации подобных пептидов.
Передача сигналов SAP интенсивно изучалась у двух видов морских ежей, Strongylocentrotus purpuratus (Darszon et al., 2011) и Arbacia punctulata (Kaupp et al., 2008), с использованием соответствующих SAP speract. и resact соответственно. Хотя общие сигнальные каскады у этих двух видов очень похожи, молекулярная идентичность лигандных рецепторов различна.Хотя рецептор resact представляет собой мембранную форму гуанилилциклазы (GC) (Singh et al., 1988), speract связывается с Cys-богатым белком 77 кДа (Dangott and Garbers, 1984; Dangott et al., 1989), который каким-то образом активирует мембранный GC. Один интересный вопрос заключается в том, как развивались различные комбинации лиганд-рецептор без изменения общего сигнального каскада у иглокожих. Следует отметить, что speract и его изоформы продуцируются в дополнительных клетках (соматических клетках) ооцита (Kinoh et al., 1994), тогда как asterosap, который напрямую связывается с GC, продуцируется в самом ооците (Matsumoto et al., 1999).
Хотя общие сигнальные каскады у этих двух видов очень похожи, молекулярная идентичность лигандных рецепторов различна.Хотя рецептор resact представляет собой мембранную форму гуанилилциклазы (GC) (Singh et al., 1988), speract связывается с Cys-богатым белком 77 кДа (Dangott and Garbers, 1984; Dangott et al., 1989), который каким-то образом активирует мембранный GC. Один интересный вопрос заключается в том, как развивались различные комбинации лиганд-рецептор без изменения общего сигнального каскада у иглокожих. Следует отметить, что speract и его изоформы продуцируются в дополнительных клетках (соматических клетках) ооцита (Kinoh et al., 1994), тогда как asterosap, который напрямую связывается с GC, продуцируется в самом ооците (Matsumoto et al., 1999).
Speract является первым идентифицированным SAP (Gly-Phe-Asp-Leu-Asn-Gly-Gly-Gly-Val-Gly), и его структурно-функциональная взаимосвязь хорошо известна благодаря существованию многих природных изоформ (Suzuki, 1995 ) и некоторые синтетические аналоги (Garbers et al. , 1982; Nomura, Isaka, 1985; Tatsu et al., 2002) этого пептида. Интересно, что наиболее важным остатком является Gly в шестом положении, поскольку его замена любой аминокислотой приводит как минимум к 1000-кратному снижению сродства к его рецептору (Tatsu et al., 2002). Для биохимических и физиологических исследований speract и его аналоги (или изоформы) были получены очисткой природных пептидов или химическим синтезом. Чтобы изучить применимость рекомбинантного speract в этом исследовании, мы подготовили три рекомбинантные версии этого пептида, слитые с флуоресцентными белками (FP), продуцируемыми в Escherichia coli в качестве флуоресцентных аналогов. Хотя эти FP-меченные speracts (FP-speracts) более чем в 20 раз больше, чем интактные speract, эти лиганды активны и обладают аффинностью связывания, сравнимой с аффинностью природного лиганда.Существование множества цветовых вариаций FP способствовало получению FP-speract с различными оптическими свойствами, что позволило нам проводить различные биоанализы с использованием флуоресцентной спектроскопии для оценки активности лиганда FP-speract.
, 1982; Nomura, Isaka, 1985; Tatsu et al., 2002) этого пептида. Интересно, что наиболее важным остатком является Gly в шестом положении, поскольку его замена любой аминокислотой приводит как минимум к 1000-кратному снижению сродства к его рецептору (Tatsu et al., 2002). Для биохимических и физиологических исследований speract и его аналоги (или изоформы) были получены очисткой природных пептидов или химическим синтезом. Чтобы изучить применимость рекомбинантного speract в этом исследовании, мы подготовили три рекомбинантные версии этого пептида, слитые с флуоресцентными белками (FP), продуцируемыми в Escherichia coli в качестве флуоресцентных аналогов. Хотя эти FP-меченные speracts (FP-speracts) более чем в 20 раз больше, чем интактные speract, эти лиганды активны и обладают аффинностью связывания, сравнимой с аффинностью природного лиганда.Существование множества цветовых вариаций FP способствовало получению FP-speract с различными оптическими свойствами, что позволило нам проводить различные биоанализы с использованием флуоресцентной спектроскопии для оценки активности лиганда FP-speract. Более того, мы определили близость сайтов связывания speract на плазматической мембране сперматозоидов, используя голубой и желтый варианты FP-speract в качестве пары FRET, что дало нам новое понимание рецептора speract.
Более того, мы определили близость сайтов связывания speract на плазматической мембране сперматозоидов, используя голубой и желтый варианты FP-speract в качестве пары FRET, что дало нам новое понимание рецептора speract.
- ASW
- искусственная морская вода
- BSA
- бычий сывороточный альбумин
- [Ca 2+ ] i
- внутриклеточный Ca 2+ csosine cyospine
- DASW
- разбавленная искусственная морская вода
- eCFP
- вариант усиленного голубого флуоресцентного белка
- FP
- флуоресцентный белок
- FP-speract
- speract, помеченный флуоресцентным белком 0 FRET, перенос энергии флуоресценции
- FRET -speract
- меченный флуоресцеином speract
- GC
- гуанилилциклаза
- IPTG
- изопропил-β-D-1-тиогалактопиранозид
- LBular
- 9151 LBular
- SAP Luria 60 iBulth 9141
- пептид, активирующий сперматозоиды
- SAP Luria 60 iBulth 9141
- sNHE 916 03 сперматозоид Na + / H + обменник
- V м
- мембранный потенциал
МАТЕРИАЛЫ И МЕТОДЫ
Материалы
Половозрелые Strongylocentrotus 9000 purpuratus (sea urpuratus) получено из Памана (Энсенада, Нижняя Калифорния, Мексика) и Lytechinus pictus (Verrill 1867) из Маринуса (Лонг-Бич, Калифорния, США). Сухие сперматозоиды собирали после внутрицеломной инъекции 0,5 моль · л -1 KCl и хранили на льду до использования. Фрагмент Кленова, HindIII, XhoI, XbaI, NcoI, лигаза Т4, набор GeneJET Plasmid Miniprep, набор для экстракции геля GeneJET, лестницы ДНК и агароза Ni-NTA Superflow были от Thermo Scientific (Waltham, MA, США). ДНК-полимераза Vent была от New England Biolabs (Ипсвич, Массачусетс, США). Citifluor был от компании Electron Microscopy Sciences (Хатфилд, Пенсильвания, США). Флуоресцентные красители BCECF-AM, 3,3′-дипропилтиадикарбоцианин йодид [DiSC 3 (5)] и Fluo-4-AM были получены от Invitrogen-Molecular Probes (Юджин, Орегон, США).Коктейльные таблетки с ингибитором протеазы cOmplete ™ были от Roche (Базель, Швейцария). Бульон Лурия (LB; содержащий триптон, дрожжевой экстракт и NaCl), канамицин и остальные реагенты были от Sigma-Aldrich. Синтетический спектр был предоставлен доктором Тацу (AIST, Ikeda, Япония), а меченный флуоресцеином спектр (F-speract) был приготовлен, как описано ранее (Nishigaki and Darszon, 2000).
Сухие сперматозоиды собирали после внутрицеломной инъекции 0,5 моль · л -1 KCl и хранили на льду до использования. Фрагмент Кленова, HindIII, XhoI, XbaI, NcoI, лигаза Т4, набор GeneJET Plasmid Miniprep, набор для экстракции геля GeneJET, лестницы ДНК и агароза Ni-NTA Superflow были от Thermo Scientific (Waltham, MA, США). ДНК-полимераза Vent была от New England Biolabs (Ипсвич, Массачусетс, США). Citifluor был от компании Electron Microscopy Sciences (Хатфилд, Пенсильвания, США). Флуоресцентные красители BCECF-AM, 3,3′-дипропилтиадикарбоцианин йодид [DiSC 3 (5)] и Fluo-4-AM были получены от Invitrogen-Molecular Probes (Юджин, Орегон, США).Коктейльные таблетки с ингибитором протеазы cOmplete ™ были от Roche (Базель, Швейцария). Бульон Лурия (LB; содержащий триптон, дрожжевой экстракт и NaCl), канамицин и остальные реагенты были от Sigma-Aldrich. Синтетический спектр был предоставлен доктором Тацу (AIST, Ikeda, Япония), а меченный флуоресцеином спектр (F-speract) был приготовлен, как описано ранее (Nishigaki and Darszon, 2000). Искусственная морская вода (ASW), содержащаяся в ммоль л -1 : 486 NaCl, 27 MgCl 2 , 29 MgSO 4 , 10 CaCl 2 , 10 KCl, 2.5 NaHCO 3 , 0,1 ЭДТА, 10 HEPES, pH 8,0 и 1000 мОсм л -1 . Low-Ca 2+ ASW содержала 1 ммоль л -1 Ca 2+ , pH 7,0. Сперматозоиды набухали, как описано в другом месте (Babcock et al., 1992; Reynaud et al., 1993) в 10-кратном разведении ASW (DASW) с добавлением 20 ммоль л -1 MgSO 4 , 120–160 мОсм, pH 6.8. Штаммы E. coli JM109 (DE3) и BL21 (DE3) были получены из Национального института биологических ресурсов , штамм E. coli , Национальный институт генетики, Мисима, Япония.Плазмиды, кодирующие eCFP-pRSETB (Kotera et al., 2010), mVenus-pRSETB (Nagai et al., 2002) и pmAmetrine1.2 (Ding et al., 2011), были предоставлены доктором T. Nagai (Университет Осаки), Dr А. Мияваки (RIKEN) и доктор Р. Кэмпбелл (Университет Альберты, плазмида Addgene № 42171), соответственно.
Искусственная морская вода (ASW), содержащаяся в ммоль л -1 : 486 NaCl, 27 MgCl 2 , 29 MgSO 4 , 10 CaCl 2 , 10 KCl, 2.5 NaHCO 3 , 0,1 ЭДТА, 10 HEPES, pH 8,0 и 1000 мОсм л -1 . Low-Ca 2+ ASW содержала 1 ммоль л -1 Ca 2+ , pH 7,0. Сперматозоиды набухали, как описано в другом месте (Babcock et al., 1992; Reynaud et al., 1993) в 10-кратном разведении ASW (DASW) с добавлением 20 ммоль л -1 MgSO 4 , 120–160 мОсм, pH 6.8. Штаммы E. coli JM109 (DE3) и BL21 (DE3) были получены из Национального института биологических ресурсов , штамм E. coli , Национальный институт генетики, Мисима, Япония.Плазмиды, кодирующие eCFP-pRSETB (Kotera et al., 2010), mVenus-pRSETB (Nagai et al., 2002) и pmAmetrine1.2 (Ding et al., 2011), были предоставлены доктором T. Nagai (Университет Осаки), Dr А. Мияваки (RIKEN) и доктор Р. Кэмпбелл (Университет Альберты, плазмида Addgene № 42171), соответственно.
Конструирование плазмид для FP-speract
Мы использовали модифицированный pET28c (Novagen) в качестве вектора экспрессии для рекомбинантных белков. Используя следующие два праймера, 5 ‘CCC TCT AGA AAT AAT TTT GTT TAA CTT TAA GA и CCC AAG CTT ATA TCT CCT TCT TAA AGT TAA AC 3’, с использованием фрагмента Кленова синтезировали двухцепочечную ДНК.Эту двухцепочечную ДНК дважды расщепляли XbaI / HindIII и вставляли в вектор pET28c с использованием тех же рестрикционных ферментов, что и сайты клонирования. Следовательно, модифицированный pET28c, названный pET28HindIII, потерял приблизительно 120 п.н. pET28c, включая NcoI в качестве сайта инициации трансляции и большинство других сайтов множественного клонирования исходного вектора.
Три плазмиды, кодирующие усиленный голубой флуоресцентный белок (eCFP), mVenus и mAmetrine (mAmetrine1.2), были отдельно использованы в качестве матрицы, и были выполнены ПЦР для получения одного фрагмента ДНК, который кодирует всю открытую рамку считывания каждого FP. -speract.Олигонуклеотидные праймеры для ПЦР следующие: прямой праймер (5 ‘CCA AGC TTA TGC ATC ATC ATC ACC ACC ACG TGA GCA AGG GCG AGG AGC TGT TC 3’) и обратный праймер (5 ‘CCC CTC GAG TTA GCC CAC ACC GCC ACC GTT CAG ATC AAA GCC GCC ACC GCC GCT ACC ACC ACC CTT GTA CAG CTC GTC CAT GCC GAG 3 ‘). Прямой праймер кодирует сайт HindIII и метку 6xHis, за которой следует N-конец этих FP. Обратный праймер кодирует сайт XhoI, стоп-кодон, speract, семь аминокислот в качестве линкера (Gly-Gly-Gly-Ser-Gly-Gly-Gly) и С-конец этих FP.Все ПЦР проводили с двумя единицами ДНК-полимеразы Vent, 0,3 мкмоль на -1 праймеров, 100 нг матрицы и 200 мкмоль на -1 каждого dNTP в 50 мкл с использованием следующего термического цикла: 3 мин при 96 ° C , 35 × (30 с при 96 ° C; 50 с при 58 ° C; 1 мин при 72 ° C) и 1 мин при 72 ° C. Каждая ПЦР давала одну полосу ДНК размером приблизительно 800 п.н. Продукты ПЦР очищали, дважды переваривали HindIII / XhoI и вставляли в вектор pET28HindIII с использованием тех же рестрикционных ферментов, что и сайты клонирования.
-speract.Олигонуклеотидные праймеры для ПЦР следующие: прямой праймер (5 ‘CCA AGC TTA TGC ATC ATC ATC ACC ACC ACG TGA GCA AGG GCG AGG AGC TGT TC 3’) и обратный праймер (5 ‘CCC CTC GAG TTA GCC CAC ACC GCC ACC GTT CAG ATC AAA GCC GCC ACC GCC GCT ACC ACC ACC CTT GTA CAG CTC GTC CAT GCC GAG 3 ‘). Прямой праймер кодирует сайт HindIII и метку 6xHis, за которой следует N-конец этих FP. Обратный праймер кодирует сайт XhoI, стоп-кодон, speract, семь аминокислот в качестве линкера (Gly-Gly-Gly-Ser-Gly-Gly-Gly) и С-конец этих FP.Все ПЦР проводили с двумя единицами ДНК-полимеразы Vent, 0,3 мкмоль на -1 праймеров, 100 нг матрицы и 200 мкмоль на -1 каждого dNTP в 50 мкл с использованием следующего термического цикла: 3 мин при 96 ° C , 35 × (30 с при 96 ° C; 50 с при 58 ° C; 1 мин при 72 ° C) и 1 мин при 72 ° C. Каждая ПЦР давала одну полосу ДНК размером приблизительно 800 п.н. Продукты ПЦР очищали, дважды переваривали HindIII / XhoI и вставляли в вектор pET28HindIII с использованием тех же рестрикционных ферментов, что и сайты клонирования. Лигированные продукты использовали для трансформации Ca 2+ -компетентных клеток JM109 (DE3). Несколько колоний инокулировали в 3 мл среды LB без изопропил-β-D-1-тиогалактопиранозида (IPTG) и инкубировали в течение ночи при 37 ° C при 200 об / мин. После центрифугирования клеточной суспензии осадки бактерий, содержащие ожидаемые плазмиды, показали соответствующие цвета каждого FP (мы подтвердили присутствие мАметрина путем возбуждения светом 405 нм, поскольку он имел слабый желтый цвет). Последовательности ДНК открытой рамки считывания каждого FP-speract были подтверждены средством секвенирования ДНК нашего института.
Лигированные продукты использовали для трансформации Ca 2+ -компетентных клеток JM109 (DE3). Несколько колоний инокулировали в 3 мл среды LB без изопропил-β-D-1-тиогалактопиранозида (IPTG) и инкубировали в течение ночи при 37 ° C при 200 об / мин. После центрифугирования клеточной суспензии осадки бактерий, содержащие ожидаемые плазмиды, показали соответствующие цвета каждого FP (мы подтвердили присутствие мАметрина путем возбуждения светом 405 нм, поскольку он имел слабый желтый цвет). Последовательности ДНК открытой рамки считывания каждого FP-speract были подтверждены средством секвенирования ДНК нашего института.
Получение FP-speract
Каждый вектор экспрессии FP-speract трансформировали в штамм E.coli BL21 (DE3). Единственную колонию выращивали в 50 мл среды LB с канамицином (30 мкг мл -1 ) в течение 12 ч при 37 ° C. Затем 2 мл переносили в 200 мл среды LB с канамицином (30 мкг мл -1 ) и инкубировали при 37 ° C. Когда OD 600 достигла 0,8, добавляли 300 мкмоль · 1 -1 IPTG и инкубацию продолжали при 30 ° C в течение 24 часов. Клетки центрифугировали (4000 г в течение 20 мин при 4 ° C) и осадок трижды промывали NaCl 150 ммоль л -1 , трис-HCl 10 ммоль л -1 , pH 7,4 ( раствор А) и повторно суспендировали с 8 мл раствора А, содержащего cOmplete ™. Клетки лизировали обработкой ультразвуком с помощью ультразвукового процессора GE50T (три обработки по 30 с при мощности 20 Вт на льду, разделенных интервалами в 1 мин) и центрифугировали (15000 г в течение 15 минут при 4 ° C). Супернатант пропускали через никелевую колонку Ni-NTA Superflow и промывали раствором А, содержащим 50 ммоль 1 -1 имидазола.Затем FP-speract элюировали раствором A, содержащим 250 ммоль -1 имидазола. Процесс очистки проводился при комнатной температуре. Концентрацию каждого FP-speract определяли с помощью анализа белка Брэдфорда с использованием бычьего сывороточного альбумина (BSA) в качестве стандарта. Для физиологических измерений ( V m , pH i и [Ca 2+ ] i ) мы использовали 10 нмоль l -1 eCFP-speract, который определяли по его абсорбции (молярная экстинкция).
Клетки центрифугировали (4000 г в течение 20 мин при 4 ° C) и осадок трижды промывали NaCl 150 ммоль л -1 , трис-HCl 10 ммоль л -1 , pH 7,4 ( раствор А) и повторно суспендировали с 8 мл раствора А, содержащего cOmplete ™. Клетки лизировали обработкой ультразвуком с помощью ультразвукового процессора GE50T (три обработки по 30 с при мощности 20 Вт на льду, разделенных интервалами в 1 мин) и центрифугировали (15000 г в течение 15 минут при 4 ° C). Супернатант пропускали через никелевую колонку Ni-NTA Superflow и промывали раствором А, содержащим 50 ммоль 1 -1 имидазола.Затем FP-speract элюировали раствором A, содержащим 250 ммоль -1 имидазола. Процесс очистки проводился при комнатной температуре. Концентрацию каждого FP-speract определяли с помощью анализа белка Брэдфорда с использованием бычьего сывороточного альбумина (BSA) в качестве стандарта. Для физиологических измерений ( V m , pH i и [Ca 2+ ] i ) мы использовали 10 нмоль l -1 eCFP-speract, который определяли по его абсорбции (молярная экстинкция). коэффициент eCFP: 32 500 M -1 см -1 ).Качество каждого очищенного FP-спектра подтверждалось электрофорезом на 15% SDS-PAGE (фиг. S1).
коэффициент eCFP: 32 500 M -1 см -1 ).Качество каждого очищенного FP-спектра подтверждалось электрофорезом на 15% SDS-PAGE (фиг. S1).
Флуорометрические определения
V m , pH i и [Ca 2+ ] i сперматозоидов S. purpuratus определяли с использованием DiSC3 (5), BCECF-AM и Fluo-4 -AM, соответственно, как сообщалось ранее (Beltrán et al., 2014). В трех случаях для стимуляции спермы было добавлено 10 нмоль -1 speract или 10 нмоль l -1 eCFP-speract, и изменение флуоресценции регистрировалось в течение 1 мин с помощью спектрофлуориметра SLM8000, модифицированного Olis Global Works. Программного обеспечения. V м Измерения и pHi проводились как в DASW, так и в обычном ASW. Чтобы нормализовать изменения [Ca 2+ ] –, клетки пермеабилизировали 0,05% Triton X-100, и максимальная флуоресценция Fluo-4 была получена в каждой записи.
Визуализация связывания mVenus-speract с рецептора сперматозоидов L. pictus
L ytechinus pictus сперматозоидов разводили в 10 раз в низко-Ca 2+ ASW, а затем в 100 раз в ASW, содержащем 0.1% BSA и 10 нмоль l -1 mVenus-speract или 10 нмоль l -1 mVenus-speract с 500 нмоль l -1 speract (в качестве конкурента). Образцы центрифугировали (1000 г в течение 7 мин при 4 ° C) и супернатант отбрасывали для удаления несвязанного белка. Сперму (25 мкл суспензии) прикрепляли к трехлуночному предметному стеклу (Electron Microscopy Sciences) при 4 ° C в течение 1 часа и фиксировали 25 мкл 4% параформальдегида в ASW в течение 10 минут. Затем клетки трижды промывали фосфатно-солевым буферным раствором (в ммоль · l -1 : 137 NaCl, 2.7 KCl, 10 Na 2 HPO 4 и 1,8 KH 2 PO 4 , pH 7,4) и 5 мкл Citifluor были добавлены для минимизации флуоресцентного обесцвечивания. Образцы закрывали покровным стеклом (толщиной 0,13–0,17 мм), а края покрывали лаком для ногтей. Флуоресценцию mVenus-speract на сперматозоидах наблюдали с объективом 100 × на конфокальном микроскопе Zeiss LSM510 с использованием аргонового лазера (488 нм) для возбуждения. Изображения спермы были получены с помощью программы LSM 5 Image Browser.
Образцы закрывали покровным стеклом (толщиной 0,13–0,17 мм), а края покрывали лаком для ногтей. Флуоресценцию mVenus-speract на сперматозоидах наблюдали с объективом 100 × на конфокальном микроскопе Zeiss LSM510 с использованием аргонового лазера (488 нм) для возбуждения. Изображения спермы были получены с помощью программы LSM 5 Image Browser.
Анализы конкурентного связывания с химически синтезированным аналогом флуоресцентного speract (F-speract)
Анализы конкурентного связывания с использованием F-speract проводили, как сообщалось ранее (Tatsu et al., 2002), с некоторыми модификациями. В этой работе мы измерили флуоресценцию 10 нмоль l -1 F-speract в суспензии сперматозоидов L. pictus , которая содержала 10 нмоль l -1 эквивалентов рецептора speract, с использованием кюветы небольшого объема. (45 мкл). Как обязательный конкурент, 1, 3.16, 10, 31,6, 100, 316 и 1000 нмоль 1 -1 либо speract, либо mAmetrine-speract добавляли к образцу и определяли их интенсивности флуоресценции (λ ex 494 / λ em 524 нм). Несмотря на то, что интенсивность флуоресценции мАметрина, возбуждаемого при длине волны 494 нм, довольно мала при эквивалентной концентрации F-speract, его флуоресценцией при гораздо более высокой концентрации нельзя пренебречь. Следовательно, чистые интенсивности флуоресценции F-speract получали вычитанием интенсивностей флуоресценции mAmetrine-speract (в суспензии сперматозоидов) из общих интенсивностей флуоресценции.
Несмотря на то, что интенсивность флуоресценции мАметрина, возбуждаемого при длине волны 494 нм, довольно мала при эквивалентной концентрации F-speract, его флуоресценцией при гораздо более высокой концентрации нельзя пренебречь. Следовательно, чистые интенсивности флуоресценции F-speract получали вычитанием интенсивностей флуоресценции mAmetrine-speract (в суспензии сперматозоидов) из общих интенсивностей флуоресценции.
FRET-анализы с использованием eCFP-speract и mVenus-speract
Суспензию сперматозоидов ( L. pictus ), содержащую 10 нмоль 1 -1 эквивалентов рецептора speract, смешивали с комбинацией eCFP-speract и mVenus-speract в наличие или отсутствие 1 мкмоль л -1 speract. Использовали два разных состава лигандов следующим образом; 5 нмоль l -1 eCFP-speract и 5 нмоль l -1 mVenus-speract для соотношения 1/1 (донор / акцептор) и 2 нмоль l -1 eCFP-speract и 8 нмоль l — 1 мVenus-speract для соотношения 1/4. Впоследствии спектр излучения (λ из = 433 нм) образца (с конкурентом и без него) регистрировали в спектрофлуориметре.
Впоследствии спектр излучения (λ из = 433 нм) образца (с конкурентом и без него) регистрировали в спектрофлуориметре.
Статистический анализ
Спектры или интенсивности флуоресценции были получены с помощью программного обеспечения 4.2 Olis Global Works и обработаны с помощью Microsoft Excel или R project 3.0.1 (бесплатное программное обеспечение). На рисунках все значения представляют собой средние значения ± s.e.m. если не указано другое. Мы провели как минимум три разных эксперимента с тремя разными морскими ежами.Тест Шапиро-Уилка показал, что все данные, полученные в этом исследовании, показали нормальное распределение ( P > 0,05). Статистическая значимость оценивалась с помощью критерия Стьюдента t . Различия считали статистически значимыми при P <0,05.
РЕЗУЛЬТАТЫ
Дизайн FP-speract
Сообщалось, что модификация аминоконца speract небольшими молекулами не влияет на его лигандную активность (Dangott and Garbers, 1984; Nishigaki and Darszon, 2000). Поэтому мы решили слить speract с FP на аминоконце этого пептидного лиганда. На рис. 1 показана схематическая структура FP-speract. Принимая во внимание, что объемный FP может мешать взаимодействиям speract-рецептор, относительно длинный спейсер, состоящий из семи неструктурных аминокислот (Gly-Gly-Gly-Ser-Gly-Gly-Gly), был вставлен между FP и speract. Чтобы облегчить процесс очистки, мы добавили 6xHis к аминоконцу FP, используя прямой праймер PCR. Как описано в разделе «Материалы и методы», три различных FP-спектра были получены в BL21 (DE3) и очищены на колонке Ni 2+ .Очищенные белки имеют ожидаемую молекулярную массу (~ 28 кДа), на что указывает 15% SDS-PAGE (фиг. S1).
Поэтому мы решили слить speract с FP на аминоконце этого пептидного лиганда. На рис. 1 показана схематическая структура FP-speract. Принимая во внимание, что объемный FP может мешать взаимодействиям speract-рецептор, относительно длинный спейсер, состоящий из семи неструктурных аминокислот (Gly-Gly-Gly-Ser-Gly-Gly-Gly), был вставлен между FP и speract. Чтобы облегчить процесс очистки, мы добавили 6xHis к аминоконцу FP, используя прямой праймер PCR. Как описано в разделе «Материалы и методы», три различных FP-спектра были получены в BL21 (DE3) и очищены на колонке Ni 2+ .Очищенные белки имеют ожидаемую молекулярную массу (~ 28 кДа), на что указывает 15% SDS-PAGE (фиг. S1).
Схематическая структура FP-speract. (A) Открытая рамка считывания ДНК, которая кодирует FP-speract (eCFP-speract). Прямой праймер (F-праймер) кодирует сайт HindIII, 6 × His и N-конец eCFP. Обратный праймер (R-праймер) кодирует сайт XhoI, speract, гибкий линкер (GGGSGGG) и С-конец eCFP. (B) Структура FP-speract (eCFP-speract), в которой трехмерная структура eCFP показана на ленточной диаграмме вместе с 6 остатками His на N-конце eCFP (коричневые эллипсы) и линкер, состоящий из из семи аминокислот (темно-зеленые эллипсы), за которыми следует сегмент на С-конце eCFP (черные эллипсы).Ленточная диаграмма была подготовлена с помощью пробной версии PyMOL (Schrödinger). Некоторые N- и C-концевые аминокислоты (MVSK и LGMDELYK соответственно) не включены в кристаллическую структуру eCFP (Lelimousin et al., 2009), поэтому эти аминокислоты были добавлены в виде гибких концевых хвостов (голубые эллипсы). mVenus-speract и mAmetrine-speract имеют в основном одинаковую структуру.
(B) Структура FP-speract (eCFP-speract), в которой трехмерная структура eCFP показана на ленточной диаграмме вместе с 6 остатками His на N-конце eCFP (коричневые эллипсы) и линкер, состоящий из из семи аминокислот (темно-зеленые эллипсы), за которыми следует сегмент на С-конце eCFP (черные эллипсы).Ленточная диаграмма была подготовлена с помощью пробной версии PyMOL (Schrödinger). Некоторые N- и C-концевые аминокислоты (MVSK и LGMDELYK соответственно) не включены в кристаллическую структуру eCFP (Lelimousin et al., 2009), поэтому эти аминокислоты были добавлены в виде гибких концевых хвостов (голубые эллипсы). mVenus-speract и mAmetrine-speract имеют в основном одинаковую структуру.
Speract и eCFP-speract вызывают аналогичные V m , pH i и [Ca 2+ ] i изменения в сперматозоидах
Чтобы оценить активность FP-speract, мы сначала определили способность eCFP-speract вызывать физиологические реакции сперматозоидов: V m изменения и увеличения pH i и [Ca 2+ ] i с использованием флуоресцентных индикаторов DiSC 3 (5), BCECF- AM и Fluo-4-AM, соответственно (Beltrán et al. , 2014). Сигналы флуоресценции 10 нмоль / л -1 eCFP (λ ex = 433 нм, λ em = 476 нм) не мешали измерениям флуоресценции этих индикаторов.
, 2014). Сигналы флуоресценции 10 нмоль / л -1 eCFP (λ ex = 433 нм, λ em = 476 нм) не мешали измерениям флуоресценции этих индикаторов.
Поскольку начальную гиперполяризацию, вызванную speract, трудно обнаружить в сперматозоидах S. purpuratus в нормальном ASW, мы измерили V m (а также pH и ) изменения набухших сперматозоидов в DASW, где легко наблюдать гиперполяризацию (Babcock et al., 1992). Наши результаты показывают, что 10 нмоль l -1 eCFP-speract и 10 нмоль l -1 speract вызывают аналогичные изменения в V m (рис. 2A). Аналогичным образом, 10 нмоль l -1 eCFP-speract вызывал увеличение pH i (фиг. 2B) и [Ca 2+ ] i (фиг. 2C), как и интактный speract. Стоит отметить, что величина увеличения pH –, вызванного speract, больше при DASW, чем наблюдаемое при нормальной ASW (рис.2Б). Учитывая, что специфический для сперматозоидов обменник Na + / H + (sNHE) активируется гиперполяризацией V m (Lee and Garbers, 1986) и что эта гиперполяризация больше в DASW (рис. 2A), усиленное увеличение pH – в DASW поддерживает участие sNHE в сигнальном каскаде speract.
2A), усиленное увеличение pH – в DASW поддерживает участие sNHE в сигнальном каскаде speract.
Speract и eCFP-speract вызывают аналогичные изменения мембранного потенциала ( V m ), внутриклеточного pH (pH i ) и внутриклеточной концентрации Ca 2+ ([Ca 2+ ] i ) в сперме. На левой панели показаны репрезентативные ответы сперматозоидов Strongylocentrotus purpuratus [(A) V m , (B) pH i и (C) [Ca 2+ ] i )] на 10 нмоль l −1 speract (черная линия) или 10 нмоль l −1 eCFP-speract (красная линия). Для измерений V m и pHi ответы сперматозоидов определяли в разбавленной ASW (DASW), а также в нормальной ASW. Стрелки указывают на добавление лиганда. F / F 0 , изменение интенсивности флуоресценции относительно исходного уровня.Правые панели показывают средние значения максимальной реакции сперматозоидов на speract (черные столбцы) или eCFP-speract (красные столбцы), за исключением гиперполяризации V m , полученной в DASW в конце измерений. Планки погрешностей представляют собой s.e.m. ( п = 3). Нет разницы между ответами сперматозоидов, вызванными speract, и ответами, вызванными eCFP-speract, ни в одном из случаев.
Планки погрешностей представляют собой s.e.m. ( п = 3). Нет разницы между ответами сперматозоидов, вызванными speract, и ответами, вызванными eCFP-speract, ни в одном из случаев.
Визуализация связывания mVenus-speract с рецептором сперматозоидов
Ранее мы подготовили F-speract как синтетический флуоресцентный аналог speract (Nishigaki and Darszon, 2000).F-speract является полезным аналогом speract, с которым можно изучать взаимодействия лиганд-рецептор, поскольку его флуоресценция гаснет при связывании с его рецептором. Однако F-speract не позволяет визуализировать его связывание с рецептором под флуоресцентной микроскопией. Напротив, mVenus-speract сохранял свою флуоресценцию при связывании рецептора и позволял нам визуализировать локализацию рецепторов speract с этим флуоресцентным лигандом (рис. 3). В присутствии избытка speract сигнал флуоресценции mVenus не наблюдался, что указывает на специфичность сигнала флуоресценции от жгутика сперматозоидов.
Рис. 3.
3.mVenus-speract специфически связывается со жгутиком. Верхние панели показывают конфокальные флуоресцентные изображения, а нижние панели представляют собой наложение флуоресцентных и фазово-контрастных изображений. На левой панели представлены репрезентативные изображения, полученные с использованием сперматозоидов Lytechinus pictus (контроль). Центральные панели показывают изображения сперматозоидов, обработанных 10 нмоль · 1 -1 мVenus-speract. Правые панели показывают изображения, полученные в присутствии избытка speract (500 нмоль l -1 ) с 10 нмоль l -1 mVenus-speract.
Конкурентная связывающая активность FP-speract с использованием F-speract в качестве флуоресцентного зонда
Хотя 10 нмоль l -1 как FP-speract, так и speract индуцировали аналогичные ответы сперматозоидов (рис. 2), необходимо было выполнить количественный анализ. эксперименты по точной оценке активности FP-speract. С этой целью мы провели анализ конкурентного связывания с использованием F-speract в качестве флуоресцентного зонда, который, как предполагается, является наиболее эффективным методом оценки лигандной активности аналогов speract (Tatsu et al. , 2002). Для этого эксперимента мы использовали сперматозоидов L. pictus , потому что количество сайтов связывания speract у этого вида в три раза выше, чем у S. purpuratus . Одной из проблем при проведении этого эксперимента является перекрытие флуоресценции между F-speract и FP-speract. Как упоминалось ранее, пик излучения флуоресценции eCFP отличается от пика флуоресцеина (F-speract). Однако высокая концентрация eCFP полностью маскировала сигнал флуоресценции F-speract из-за значительного перекрытия флуоресценции между флуоресцеином и eCFP.Впоследствии мы попытались использовать для этого эксперимента синий FP (mEBFP2)-тегированный speract, но нам не удалось произвести достаточное количество растворимого mEBFP2-speract в BL21 (DE3) по неизвестной причине. Таким образом, мы использовали мАметрин в качестве полезной альтернативы для этой цели. мАметрин представляет собой желтый FP (λ em = 526 нм) с пиком возбуждения около 400 нм (Ai et al., 2008; Ding et al. , 2011), который классифицируется как флуорофор с большим стоксовым сдвигом (120 нм между пики возбуждения и излучения). Несмотря на то, что испускание флуоресценции мАметрина при возбуждении на длине волны 490 нм незначительно по сравнению с эмиссией, полученной при возбуждении на длине волны 400 нм при сопоставимой концентрации F-speract, этой небольшой интенсивностью флуоресценции нельзя пренебречь при более высоких концентрациях флуорофора.Поэтому для этого эксперимента мы вычитали интенсивность флуоресценции самого мАметрина из общей интенсивности флуоресценции, чтобы получить чистое тушение F-speract. На фиг.4 показаны кривые конкурентного связывания mAmetrine-speract и speract при связывании F-speract с его рецептором в сперматозоидах L. pictus . Значения IC 50 mAmetrine-speract и speract составляли 61 ± 9 и 8 ± 0,8 нмоль л -1 ( n = 3), соответственно, что указывает на то, что этот флуоресцентный аналог speract имеет 7.Сродство к рецептору в 6 раз меньше, чем у speract.
, 2002). Для этого эксперимента мы использовали сперматозоидов L. pictus , потому что количество сайтов связывания speract у этого вида в три раза выше, чем у S. purpuratus . Одной из проблем при проведении этого эксперимента является перекрытие флуоресценции между F-speract и FP-speract. Как упоминалось ранее, пик излучения флуоресценции eCFP отличается от пика флуоресцеина (F-speract). Однако высокая концентрация eCFP полностью маскировала сигнал флуоресценции F-speract из-за значительного перекрытия флуоресценции между флуоресцеином и eCFP.Впоследствии мы попытались использовать для этого эксперимента синий FP (mEBFP2)-тегированный speract, но нам не удалось произвести достаточное количество растворимого mEBFP2-speract в BL21 (DE3) по неизвестной причине. Таким образом, мы использовали мАметрин в качестве полезной альтернативы для этой цели. мАметрин представляет собой желтый FP (λ em = 526 нм) с пиком возбуждения около 400 нм (Ai et al., 2008; Ding et al. , 2011), который классифицируется как флуорофор с большим стоксовым сдвигом (120 нм между пики возбуждения и излучения). Несмотря на то, что испускание флуоресценции мАметрина при возбуждении на длине волны 490 нм незначительно по сравнению с эмиссией, полученной при возбуждении на длине волны 400 нм при сопоставимой концентрации F-speract, этой небольшой интенсивностью флуоресценции нельзя пренебречь при более высоких концентрациях флуорофора.Поэтому для этого эксперимента мы вычитали интенсивность флуоресценции самого мАметрина из общей интенсивности флуоресценции, чтобы получить чистое тушение F-speract. На фиг.4 показаны кривые конкурентного связывания mAmetrine-speract и speract при связывании F-speract с его рецептором в сперматозоидах L. pictus . Значения IC 50 mAmetrine-speract и speract составляли 61 ± 9 и 8 ± 0,8 нмоль л -1 ( n = 3), соответственно, что указывает на то, что этот флуоресцентный аналог speract имеет 7.Сродство к рецептору в 6 раз меньше, чем у speract.
mAmetrine-speract и speract имеют сравнимое сродство к рецептору в сперматозоидах Lytechinus pictus . Восстановление интенсивности флуоресценции F-speract из состояния гашения за счет конкуренции рецепторов с использованием speract (черный) и mAmetrine-speract (красный) в различных концентрациях. Планки погрешностей представляют ± s.e.m. ( п = 3). FAU, условные единицы флуоресценции.
Измерения FRET между eCFP-speract и mVenus-speract при связывании с рецептором
Как упоминалось во введении, рецептор speract представляет собой белок 77 кДа, который обладает одним трансмембранным сегментом.Хотя он имеет некоторую гомологию последовательности с богатым Cys белком-скавенджером, обнаруженным у млекопитающих, его биохимические свойства подробно не изучены (Dangott et al., 1989). Чтобы выяснить, образуют ли рецепторы speract олигомер или существуют в виде мономера в сперме морского ежа, мы провели эксперимент по связыванию на основе FRET. На рис. 5 показаны спектры излучения флуоресценции (λ ex = 433 нм) смеси 2 нмоль l -1 eCFP-speract и 8 нмоль l -1 мVenus-speract (соотношение 1/4) с Л.pictus сперматозоидов (эквивалент 10 нмоль 1 -1 рецепторов speract) в присутствии или отсутствии избыточного количества speract. Мы наблюдали значительную разницу (15,1 ± 2,0%, n = 3) интенсивностей флуоресценции CFP между двумя условиями (фиг. 5A). В нормализованных спектрах флуоресценции (рис. 5B) интенсивности флуоресценции около 528 нм, которые соответствуют пику флуоресценции mVenus (Nagai et al., 2002), выше в отсутствие конкурента, чем в присутствии.Эти результаты показывают, что между этими двумя аналогами speract существует FRET, когда они связаны с рецептором, предполагая, что рецепторы speract существуют как кластер, образующий олигомер, по крайней мере, димер, на плазматической мембране сперматозоидов. Альтернативно, один рецепторный белок speract имеет несколько сайтов связывания для этого лиганда.
На рис. 5 показаны спектры излучения флуоресценции (λ ex = 433 нм) смеси 2 нмоль l -1 eCFP-speract и 8 нмоль l -1 мVenus-speract (соотношение 1/4) с Л.pictus сперматозоидов (эквивалент 10 нмоль 1 -1 рецепторов speract) в присутствии или отсутствии избыточного количества speract. Мы наблюдали значительную разницу (15,1 ± 2,0%, n = 3) интенсивностей флуоресценции CFP между двумя условиями (фиг. 5A). В нормализованных спектрах флуоресценции (рис. 5B) интенсивности флуоресценции около 528 нм, которые соответствуют пику флуоресценции mVenus (Nagai et al., 2002), выше в отсутствие конкурента, чем в присутствии.Эти результаты показывают, что между этими двумя аналогами speract существует FRET, когда они связаны с рецептором, предполагая, что рецепторы speract существуют как кластер, образующий олигомер, по крайней мере, димер, на плазматической мембране сперматозоидов. Альтернативно, один рецепторный белок speract имеет несколько сайтов связывания для этого лиганда.
Положительный сигнал FRET наблюдается при связывании двухцветного (голубого и желтого) лиганда с рецептором speract. (A) Типичные спектры флуоресценции смеси 2 нмоль l -1 eCFP-speract и 8 нмоль l -1 мVenus-speract в суспензии сперматозоидов Lytechinus pictus (эквивалент 10 нмоль l -1 speract рецептор) в присутствии (красная линия) и отсутствии (черная линия) избытка speract (1 мкмоль л -1 ) в качестве конкурента.FAU, условные единицы флуоресценции. Двунаправленная стрелка указывает пик флуоресценции eCFP. (B) Нормализованные спектры флуоресценции для двух условий. Эти два спектра не идентичны из-за наличия FRET; Спектры флуоресценции около 528 нм (пик флуоресценции mVenus, обозначен стрелкой) без конкурента немного выше, чем в присутствии конкурента.
ОБСУЖДЕНИЕ
Разработка FP-speract
В этом исследовании мы приготовили рекомбинантный аналог speract, помеченный FP (FP-speract, рис. 1) и оценил его функциональность. Несмотря на относительно большой размер FP (25 кДа) по сравнению со speract (декапептидом), FP-speract индуцировал физиологические ответы сперматозоидов ( V m , pH i и [Ca 2+ ] i ) аналогично speract (рис. 2). Кривые конкурентного связывания FP-speract и speract (рис.4) показывают, что FP-speract сохранял сравнимое сродство к рецептору (только в 7,6 раз менее сильное, чем интактный speract), подтверждая, что FP-speract действительно является полезным лигандом для сперматозоиды.Неструктурный спейсер из семи аминокислот между FP и speract, вероятно, способствовал этому благоприятному результату. Насколько нам известно, существует несколько случаев функциональных FP-меченных пептидных лигандов, таких как паратироидный гормон и брадикинин (Charest-Morin et al., 2013). Хотя оба являются функциональными лигандами и вызывают ожидаемые клеточные ответы, точное сродство к соответствующему рецептору не определено.
1) и оценил его функциональность. Несмотря на относительно большой размер FP (25 кДа) по сравнению со speract (декапептидом), FP-speract индуцировал физиологические ответы сперматозоидов ( V m , pH i и [Ca 2+ ] i ) аналогично speract (рис. 2). Кривые конкурентного связывания FP-speract и speract (рис.4) показывают, что FP-speract сохранял сравнимое сродство к рецептору (только в 7,6 раз менее сильное, чем интактный speract), подтверждая, что FP-speract действительно является полезным лигандом для сперматозоиды.Неструктурный спейсер из семи аминокислот между FP и speract, вероятно, способствовал этому благоприятному результату. Насколько нам известно, существует несколько случаев функциональных FP-меченных пептидных лигандов, таких как паратироидный гормон и брадикинин (Charest-Morin et al., 2013). Хотя оба являются функциональными лигандами и вызывают ожидаемые клеточные ответы, точное сродство к соответствующему рецептору не определено.
Применение FP-speract для определения образования кластеров рецептором speract
Ранее мы приготовили флуоресцентные аналоги speract, меченные флуоресцеином (F-speract), флуоресценция которого гасится при связывании с его рецептором (Nishigaki and Darszon, 2000) .Напротив, FP-speract сохраняет свою флуоресценцию даже после связывания со своим рецептором (рис. 3). Воспользовавшись этим свойством, мы применили два цветовых варианта FP-speract, голубой и желтый, чтобы изучить близость сайтов связывания speract.
Рецепторы resact и asterosap известны как мембраносвязанные GC (Matsumoto et al., 2003; Singh et al., 1988). Ранее было подсчитано, что в жгутике сперматозоидов морских звезд имеется около 100000 сайтов связывания звездообразных звездочек (Nishigaki et al., 2000). Недавно сообщалось, что одна клетка из сперматозоидов A. punctulata содержит 300000 молекул ГК (Pichlo et al., 2014). Сравнивая эти числа, можно заметить, что рецепторов speract меньше: 60 000 сайтов связывания в L. pictus и 20 000 сайтов связывания в S. purpuratus (Nishigaki et al., 2001). Рецептор speract представляет собой богатый Cys белок 77 кДа, отличный от GC, который, как предполагается, связан с GC (Dangott and Garbers, 1984; Dangott et al., 1989).Известно, что рецептор предсердного натрийуретического пептида, мембраносвязанной гуанилилциклазы A, существует в виде димера (Chinkers and Wilson, 1992; Ogawa et al., 2004). Однако нет такой информации для рецепторов SAP ни в рецепторах типа resact (GC), ни в рецепторах speract-типа.
В этом исследовании мы исследовали близость между сайтами связывания speract на плазматической мембране сперматозоидов, используя многоцветный FP-speract. Обычная оптическая микроскопия не может определить расстояние между двумя объектами, локализованными на расстоянии менее 200 нм.Напротив, современные методы микроскопии сверхвысокого разрешения могут увеличить разрешение до 20 нм (Fernández-Suárez and Ting, 2008; Huang et al., 2009). Однако эти методы требуют сложных экспериментальных условий и анализа. Поэтому мы провели измерение FRET с использованием голубого и желтого вариантов FP-speract. На практике мы можем получить более высокое пространственное разрешение с помощью FRET (<10 нм), чем с помощью микроскопии сверхвысокого разрешения (Fernández-Suárez and Ting, 2008; Huang et al., 2009). Сообщалось, что расстояние Ферстера (50% FRET) между eCFP и mVenus оценивается примерно в 5 нм (Patterson et al., 2000). Поскольку эффективность FRET уменьшается в зависимости от шестой степени расстояния, удвоенное расстояние Ферстера (10 нм в этом исследовании) должно привести к эффективности FRET менее 2%. В этой работе мы получили примерно 15% эффективности FRET в наших экспериментальных условиях (соотношение донор / акцептор 1/4). Когда мы провели тот же эксперимент с соотношением 1/1, эффективность FRET была ниже (10.1 ± 0,8%, n = 4), вероятно, потому, что в этом состоянии донорским флуорофором была занята большая часть соседних сайтов связывания. Хотя рецепторы speract в большом количестве присутствуют в плазматической мембране жгутиков сперматозоидов L. pictus (60000 сайтов связывания на клетку), среднее расстояние между сайтами связывания оценивается примерно в 40 нм, если сайты связывания равномерно распределены в плазме. мембрана жгутика сперматозоидов (диаметр 0,3 мкм и длина 45 мкм).Таким образом, наши результаты (рис. 5) предполагают, что сайты связывания speract не распределены однородно, а существуют в виде кластеров. Это первое экспериментальное свидетельство кластерообразования рецепторов SAP. Учитывая, что рецептор speract содержит четыре повтора Cys-богатых доменов (Dangott et al., 1989), мы не можем исключить возможность того, что один рецептор полипептида имеет несколько сайтов связывания speract. Дальнейшие эксперименты, такие как точное определение молекулярной стехиометрии между speract рецептором и speract сайтов связывания, необходимы для решения этого вопроса.Сообщалось, что рецептор speract локализован в липидном рафте (Loza-Huerta et al., 2013; Ohta et al., 2000), что может частично объяснять его формирование кластеров. Независимо от типа кластера (межмолекулярный, внутримолекулярный или оба), кластер сайтов связывания может обеспечивать кооперативные связывания, как сообщается в speract и других SAP (Nishigaki et al., 2000, 2001; Pichlo et al., 2014).
FP-speract как инструмент для экспериментального обучения
Морской еж был использован в качестве классической превосходной модели для наблюдения за процессом оплодотворения в области репродуктивной биологии и биологии развития.Хотя документировать хемотаксис сперматозоидов у некоторых видов нетривиально, довольно легко наблюдать активность speract (SAP в целом), восстанавливая подавленную подвижность сперматозоидов в слегка подкисленной морской воде (см. Фильм 1) (Ohtake, 1976). Учитывая, что приготовление рекомбинантного белка, экспрессированного в E. coli , является стандартным протоколом в молекулярной биологии, FP-speract может быть весьма полезен для экспериментального обучения. Окрашивание FP-speract облегчает визуализацию модифицированного speract на протяжении всего препарата, а также определение его концентрации белка.
Выводы
В этом исследовании мы разработали рекомбинантные аналоги speract, помеченные FP (FP-speract). Хотя размер флуоресцентной метки более чем в 20 раз больше, чем у этого лиганда, FP-speract сохраняет активность, сравнимую с активностью speract, что указывает на то, что FP-speract является полезным инструментом для изучения действия этого пептида на сперматозоиды. Используя голубые и желтые варианты FP-speract, мы смогли определить кластерное образование сайтов связывания speract, что может указывать на некоторые роли для положительной и / или отрицательной кооперативности во взаимодействиях speract-рецептор.Мы считаем, что FP-speract — полезный инструмент не только для научных исследований, но и для экспериментального обучения. Кроме того, рекомбинантный пептид, меченный FP, обычно может быть использован для других типов пептидного лиганда.
Благодарности
Мы благодарим доктора Т. Нагаи (Университет Осаки), доктора А. Мияваки (RIKEN) и доктора Р. Кэмпбелла (Университет Альберты) за предоставление eCFP-pRSETB, mVenus-pRSETB и pmAmetrine1.2, соответственно, и Д-ру Y. Tatsu (AIST, Ikeda) за предоставление синтетического препарата.Мы благодарны Национальному институту генетики (Мисима, Япония) за предоставление двух штаммов E. coli , JM109 (DE3) и BL21 (DE3). Мы благодарим T. Olamendi (IBT-UNAM) за ВЭЖХ-анализ F-speract, X. Alvarado (IBT-UNAM) за конфокальные изображения, а также E. López, S. Becerra, J Yañez и P. Gaytán (IBT-UNAM) ) для синтеза олигонуклеотидов и секвенирования ДНК плазмид. Благодарим A. Blancas (IBT-UNAM) за заботу о морских ежах. Мы ценим критическое прочтение рукописи доктором А. Дарсзоном.Мы благодарим T. Liévano за рецензирование английской грамматики рукописи. Мы также благодарим анонимного рецензента за предложение улучшить наш эксперимент FRET.
СНОСКИ
Конкурирующие интересы
Авторы заявляют об отсутствии конкурирующих или финансовых интересов.
Вклад авторов
C.A.-H. выполнили почти все эксперименты и подготовили все рисунки, Ф. подготовили плазмиду, кодирующую eCFP-speract, Y.С.-Г. контролировал подготовку плазмид, кодирующих mVenus-speract и mAmetrine-speract, и все рекомбинантные белки, C.B. контролировал определение всех физиологических ответов сперматозоидов и визуализацию сайта связывания speract, а T.N. разработал весь проект и написал рукопись. Все авторы участвовали в интерпретации результатов и рецензировали рукопись.
Финансирование
Эта работа была поддержана Генеральным директором Асунтос дель Персональной Академической — Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (DGAPA-PAPIIT) — UNAM [IN204112 to C.B. и IN203513, T.N.] и Consejo Nacional de Ciencia y Tecnología (CONACyT) [128566 и 177138, T.N.]. F.R. и Ю.С.-Г. являются аспирантами, поддерживаемыми КОНАСИТ.
Дополнительная информация
Дополнительная информация доступна в Интернете по адресу http://jeb.biologies.org/lookup/suppl/doi:10.1242/jeb.127662/-/DC1
- Поступила 1 июля 2015 г.
- Принято 20 ноября 2015 г.
- © 2016. Издано ООО «Компания биологов»
— интонация электрогитары плоская выше 12-го лада, обычными методами не повезло
Прежде, чем мы начнем : Следует отметить, что все струны должны начинаться с одинаковой высоты на головках колышков.В последнее время у меня возникла проблема с интонацией, и оказалось, что моя струна B не начинается на той же высоте, что и другие струны на головке колышка.
Учитывая то, как струна идет остро в некоторых местах и плоско в других, этот случай сильно отличается от проблемы смены калибра, седла или регулировки фермы.
Вот почему :
Высота тона / играемой ноты в основном (есть и другие факторы, но они являются наиболее важными) зависит от ее длины от удерживаемого лада и моста (так называемая эффективная длина), натяжения струны и плотности струны (масса струны на Длина, а не объем).Не включайте «Напряжение и плотность», потому что они зависят от струны, и вы пробовали разные струны, как говорите в вопросе.
Теперь длина. Обычно интонация становится более резкой или более плоской. Этот факт станет очевидным, если вы поймете, что либо начальная длина (открытая нота) больше или меньше той, для которой предназначена гитара, либо струна расположена под углом к ладу, отличным от того, для которого она предназначена, поэтому каждый лад соответствует к различным точкам струны, отличным от того, для чего предназначена гитара.
Зная внутреннюю работу, ваша гитара в некоторых точках становится плоской, а в других случаях это означает, что это необычный случай, а не начальная длина (седельный чехол), а не угол (корпус анкерного стержня). Скорее всего сама гриф может повлиять на погоду, поменял профиль.
Отвечая на ваш вопрос. Точно ничего нельзя сказать . Лучше всего отдать мастеру, как Тим говорит в комментариях, и может случиться так, что мастер предложит перепрофилировать гриф .Перепрофилирование — это, возможно, немного с учетом ситуации с COVID-19, но в остальном это не очень редкая и очень тревожная работа. Многие пользователи здесь, в StackExchange, тоже изменили свой профиль.